Физиологические периоды развития половой системы мальчиков (обзор)
ID: 2014-01-1276-R-3332
Обзор
ГБОУ ВПО Саратовский ГМУ им. В.И. Разумовского Минздрава России, НИИ фундаментальной и клинической уронефрологии
Резюме
.
Ключевые слова
половая система, мальчики
Обзор
Роль гормонов в процессе внутриутробного формирования половой системы у плода мужского пола
Формирование половой системы начинается с 3-й недели эмбрионального развития. От мезонефроса отделяются первичные бипотенциальные гонады, парамезонефральные (Мюллеровы) и мезонефральные (Вольфовы) протоки – предшественники наружных и внутренних гениталий. C4-й недели эмбриогенеза начинается миграция герминативных клеток в первичную гонаду. Под контролем специфических генов – SRY и SOX 9 – происходит формирование тестикула из бипотенциальной гонады, дифференцировка клеток Сертоли и Лейдига.
Следующий этап внутриутробного развития обусловлен гормональной активностью эмбрионального тестикула.

Рис. 1. Схема гормональной активности эмбрионального тестикула
Антимюллеров гормон (АМГ), секретируемый фетальными клетками Сертоли, приводит к регрессии Мюллеровых протоков. Тестостерон, секретируемый фетальными клетками Лейдига, действует через андрогеновый рецептор Вольфовых протоков, индуцируя формирование придатков, семявыносящих протоков, семенных пузырьков. Под действием эмбриональной 5-альфа-редуктазы 2 тестостерон трансформируется в дигидротестостерон (ДГТ), который активирует андрогеновый рецептор предстательной железы и наружных гениталий, запускает процесс их маскулинизации.
С 10 - 12 недели внутриутробного развития начинается миграция эмбрионального тестикула от места первичного расположения в мошонку. В настоящее время доказано, что основная роль в этом процессе принадлежит гормональным факторам. Инсулиноподобный фактор роста 3 (ИФР 3), продуцируемый фетальными клетками Лейдига наряду с тестостероном, и антимюллеров гормон, синтезируемый в клетках Сертоли [IvellR., 2003], играют ведущую роль в трансабдоминальной миграции яичка. Андрогены регулируют прохождение яичка через паховый канал до дна мошонки. Секреция этих гормонов регулируется гонадотропинами, о чем свидетельствуют исследования, показывающие увеличение соотношения ЛГ/ тестостерон у мальчиков с крипторхизмом в возрасте 3-х месяцев [SuomiA.etal, 2006.].
Любые нарушения секреции или действия андрогенов могут привести к крипторхизму, часто ассоциированному с гипоспадией, расщеплением мошонки, микропенисом. Кроме того, доказано значение дефицита ЛГ в генезе паховой задержки яичка. В экспериментальном изучении гипогонадотропного гипогонадизма, обусловленного нарушением синтеза и действия гонадотропин-рилизинг гормона, показано нарушение пахово-мошоночной фазы опускания яичка. По данным клинического исследования, частота крипторхизма при полной форме гипогонадотропного гипогонадизма составила 40%, из которых 23% пациентов имели двусторонний крипторхизм, 17% – односторонний [Pitteloud, 2002].
Периоды физиологической активности гипоталамо-гипофизарно-гонадной системы
Итак, внутриутробное развитие плода – период высокой активности половых желез, результатом которой является дифференцировка наружных и внутренних гениталий. Гипоталамо-гипофизарно-гонадные взаимосвязи также формируются в этот период, они способствуют развитию гонад, регулируют процесс опускания яичек в мошонку. Совокупность генетических и гормональных факторов обеспечивает полноценное формирование половой системы к рождению ребенка.
К моменту рождения уровень половых гормонов быстро снижается, а с 6-7 дня постнатального развития вновь происходит активация гонадотропной секреции. Транзиторная активация гипоталамо-гипофизарной системы и половых желез в постнатальном периоде получила название мини-пубертат. Визуальный осмотр наружных гениталий, пальпация и орхиометрия, проводимые у мальчика в этот период, не выявляют существенных изменений. Однако использование точных методов оценки позволяет определить увеличение объёма гонад [MainK.M. etal, 2006]. Лабораторные исследования определяют повышение гонадотропинов, половых стероидов, антимюллерова гормона и ингибина Б в сыворотке крови новорожденных мальчиков со второй недели жизни до третьего - шестого месяца постнатального развития [GrinsponR.P., ReyR.A., 2010]. Морфологические исследования тестикула обнаруживают пролиферацию и созревание клеток Сертоли, специфические изменения клеток Лейдига [ZivkovicD., HadziselimovicF., 2009]. Этот период имеет крайне важное значение для дальнейшего становления репродуктивной системы. Гормональное обследование мальчиков в возрасте 1 – 3 месяцев после рождения делает возможным прогнозирование пубертата и репродуктивной функции. Имеются данные об успешной заместительной терапии препаратами рекомбинантных гонадотропинов в этот период, позволяющей улучшить потенциал фертильности пациентов с врожденным гипогонадотропным гипогонадизмом [Bougnères P. et al., 2008].
Период транзиторной постнатальной активности гипоталамо-гипофизарно-гонадной системы сменяется ювенильной паузой – периодом физиологического покоя половых желез. В это время уровень половых гормонов резко снижается и имеет допубертатные значения до наступления полового созревания. Исключение составляет антимюллеровый гормон, его уровень остаётся высоким в течение всего допубертатного периода и является маркером функционального созревания клеток Сертоли [GrinsponR.P., ReyR.A., 2010]. Вместе с тем незначительное и постепенное увеличение секреции гормонов гипофиза и гонад в этот период имеет место, что косвенно подтверждает созревание диэнцефальных структур головного мозга. Развития половых желез в этот период не происходит, оно тормозится гипофизарным гонадотропин-ингибирующим фактором, секреция которого регулируется гипоталамическими и эпифизарными нейропептидами. Гонадотропин-ингибирующий фактор очень похож на гонадотропный гормон по строению молекулы, а потому легко и прочно соединяется с рецепторами тех клеток, которые настроены на чувствительность к гонадотропинам. Никакого стимулирующего действия на половые железы он не оказывает, при этом перекрывает доступ к рецепторам гонадотропного гормона.
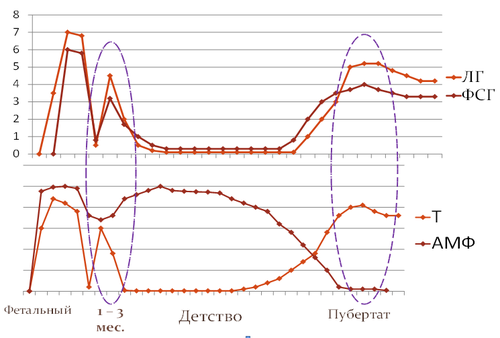
Рис.2. Схема изменения активности гипоталамо-гипофизарно-гонадной системы у мальчиков
Следующим периодом активации гипоталамо-гипофизарно-гонадной системы является пубертат. Пубертат – транзиторный период между детством и половой зрелостью, контролируется комплексом нейроэндокринных факторов, обеспечивающих ряд физических и психических изменений, приводящих к физической, психической и репродуктивной зрелости организма. Начало пубертата определяется способностью центральной нервной системы индуцировать синтез и импульсную секрецию гонадотропин-рилизинг гормона (ГТ-РГ). Результатом пульсирующей секреции ГТ-РГ является увеличение секреции гонадотропинов и стимуляция синтеза половых стероидов. Одновременно с этим повышается чувствительность гипофиза к действию гонадотропин-рилизинг гормона, происходит созревание рецепторного аппарата гонад. Лютеотропный гормон активно стимулирует синтез андрогенов в клетках Лейдига, под действием тестостерона и дигидротестостерона происходит развитие вторичных половых признаков. Фолликулостимулирующий гормон связывается с рецепторами клеток Сертоли, стимулируя продукцию ингибина Б. Уровень антимюллерова гормона к началу пубертата снижается. Под действием ФСГ в период пубертата происходит увеличение массы семенных канальцев и созревание сперматозоидов в клетках Сертоли.
Пубертатный период характеризуется становлением гипоталамо-гипофизарно-гонадных взаимосвязей, обеспечивающих работу половых желез по принципу отрицательной обратной связи. Снижение концентрации гонадотропинов на фоне пубертата является результатом обратного ингибирующего влияния половых стероидов. Учитывая особенности регуляции гонадотропной секреции, однократное исследование базального уровня гонадотропинов, как правило, неинформативно. Динамический анализ образцов сыворотки крови имеет большее диагностическое значение в определении инициации и прогрессирования пубертата.
Синтез андрогенов в надпочечниках и гонадах
По биохимической классификации, соответствующей количеству атомов углерода в молекуле, андрогены являются С-19 стероидами [А.Д. Подтетнев, 2000]. Единым субстратом для биосинтеза всех стероидных гормонов в надпочечниках и гонадах является холестерин. Андрогены синтезируются из эфиров холестерина, образующихся в печени и поступающих в эндокринные железы в составе липопротеинов низкой плотности. Внутриклеточный транспорт холестерина к внутренней мембране митохондрий регулируется преимущественно стероидогенным регуляторным протеином StAR. Биосинтез стероидов осуществляется при участии ферментов двух групп: гидроксилазы/десмолазы и дегидрогеназы/изомеразы. Гидроксилазы входят в состав цитохром Р 450- зависимой группы оксидаз. Синтез ферментов стероидогенеза в надпочечниках регулируется АКТГ, в тестикулах – ЛГ. Схема биосинтеза андрогенов представлена на рисунке 3.

Рис.3. Схема биосинтеза андрогенов в надпочечниках и гонадах
Первые три энзим-зависимых превращения являются общими для надпочечников и гонад и реализуются при участии ферментов 20,22 – холестерол-десмолазы, 17-α-гидроксилазы, 17,20-лиазы и 3-β-гидроксистероиддегидрогеназы. Заключительный этап биосинтеза половых стероидов происходит исключительно в клетках Лейдига при участии 17-β-гидроксистероиддегидрогеназы, катализирующей превращение андростендиола в тестостерон, и периферических тканях при участии 5-α-редуктазы, катализирующей синтез дигидротестостерона [И.И. Дедов, Т.В. Семичева, 2002].
Биологическое действие андрогенов в период препубертата и пубертата
Андростендион и дегидроэпиандростерон (ДГЭА) синтезируются в тестикулах в незначительном количестве. Основной пул этих андрогенов является результатом надпочечникового стероидгенеза. Андростендион и ДГЭА являются промежуточными продуктами биосинтеза тестостерона, поэтому повышение их концентрации в сыворотке крови может быть результатом дефицита фермента одного из звеньев этой цепи. Уровень дегидроэпиандростерона существенно возрастает за два года до повышения тестостерона в сыворотке. Концентрация андростендиона в крови постепенно растёт, начиная примерно с 7 лет жизни, а после 30 лет плавно снижается.
Адренархе
Началу пубертата предшествует адренархе – период препубертатной активации надпочечников, характеризующийся значительным повышением продукции надпочечниковых андрогенов. Впервые повышение уровня надпочечниковых андрогенов было установлено в 1943 г. Натаном Талботом. Ученый измерил экскрецию 17-кетостероидов с мочой у здоровых детей препубертатного возраста и обнаружил их увеличение [TalbotN.B., 1943]. Фаллер Олбрайт и его последователи определили, что появление аксилярного и лобкового оволосения у пациентов с дисгенезией гонад связано с секрецией надпочечниковых андрогенов [AlbrightF., 1942]. Ф. Олбрайт назвал этот процесс адренархе. Наступление адренархе соответствует возрасту 6 - 9 лет – приблизительно за 2 года до начала созревания гонад и манифестации физиологического пубертата. Маркером адренархе является увеличение продукции дегидроэпиандростерона и ДГЭА-сульфата, синтезирующихся в сетчатой зоне коры надпочечников. По результатам нескольких клинических исследований, уровень ДГЭА-сульфата более 40-50 мкг/дл соответствует наступлению адренархе [Kelnar C.J.H., Brook C.G.D.,1983; Wierman M.E.etal., 1986]. Регуляция адренархе недостаточно изучена, но констатирована гиперплазия сетчатой зоны коры надпочечников и повышение активности ферментов стероидогенеза: цитохрома Р450 с17 (CYP 17), ДГА-сульфтрансферазы (SULT2A1), 3β-гидроксистероиддегидрогеназы 2 типа (HSD3B2). Изменение соотношения андрогенов в сыворотке крови является следствием нарушения функции соответствующих ферментов стероидогенеза [NakamuraY., 2009].
Биологическая роль адренархе
В двух независимых исследованиях проведена оценка роста у здоровых детей в период адренархе. Небольшой, но значимый скачок роста был обнаружен в возрасте от 6,5 до 8,5 лет [Zemel B.S., Katz S.H., 1986; Largo R.H.,1993]. Другими авторами показана положительная корреляция показателей костного возраста и ДГЭА у пациентов с преждевременным половым развитием на фоне лечения гонадотропин-рилизинг-гормоном и подавления секреции гонадотропинов [Wierman M.E., 1986].
В проспективном исследовании, проводимом на большой когорте здоровых детей [OngK.K., PotauN., 2004], была обнаружена взаимосвязь между антропометрическими показателями в постнатальном периоде, избыточной прибавкой массы тела в течение препубертатного периода и высоким уровнем надпочечниковых андрогенов (дегидроэпиандростерона сульфата, андростендиона) в возрасте 8 лет для детей обоих полов. Полученные результаты показали, что исследование секреции андрогенов у детей препубертатного возраста имеет важное значение для определения риска возникновения инсулинорезистентности и центрального ожирения у взрослых.
Развитие лобкового оволосения у мальчиков, cответствующее стадиям P1 – P5 по Таннеру, прямо коррелирует с уровнем дегидроэпиандростерона, 17-β-тестостерона, 5-α-дигидротестостерона. Надпочечниковые андрогены активируют созревание гипоталамо-гипофизарно-гонадной оси, инициируют центральные механизмы пубертата. Кроме того, они снижают уровень секс-стероид-связывающего глобулина (СССГ), тем самым могут влиять на темп полового созревания за счет увеличения концентрации свободного тестостерона [LourdesI.N., 2000].
Тестостерон является основным продуктом секреции яичек в результате их стимуляции ЛГ. Большая часть тестостерона (20-78%), поступающего в кровь, связывается со специфическим транспортным белком – глобулином, связывающим половые гормоны (ГСПГ). Эта связь непрочная, тестостерон легко высвобождается, превращаясь в активную форму, и оказывает биологическое воздействие на андрогенные рецепторы тканей. Физиологически высокий уровень ГСПГ сохраняется у мальчиков в течение всего допубертатного периода, а к началу полового созревания его уровень резко снижается [SorensenK., AndersonA.M., 2007].
Тестостерон, доминирующий среди циркулирующих андрогенов, является одновременно активным гормоном и прогормоном для синтеза 5α-дигидротестостерона. Тестостерон диссоциирует от ГСПП и диффундирует в клетку. Конверсия тестостерона в дигидротестостерон происходит непосредственно в андроген-зависимых тканях и регулируется двумя изоэнзимами: 5-α-редуктазой I и II типа. Известно, что основное биологическое действие 5α-дигидротестостерона проявляется внутриутробно в развитии наружных половых органов плода мужского пола. В период пубертата ДГТ несет основную ответственность за рост и дифференцировку предстательной железы [Wilson J.D., 1996]. Методом иммуногистохимии ферментативная активность 5α-RD2 была определена и в других урогенитальных тканях: придатках, яичках, губернакулюме, кавернозных телах. Мутация гена 5αRD2 приводит к редкому аутосомно-рецессивному варианту нарушения формирования пола. Подавление активности данного фермента – к регрессии предстательной железы [Zhu Y.S., Imperato-McGinley J.L., 2009]. Экспрессия 5α-RD 1 типа происходит в печени, коже, эпителиальных клетках придатков.
Андрогены осуществляют своё действие посредством связывания с внутриклеточным андрогеновым рецептором. Взаимодействие андрогена с рецептором индуцирует ряд конформационных изменений, таких как димеризация, внутриклеточный транспорт и связывание со специфической последовательностью ДНК, целью которой является транскрипция гена – синтез РНК [QuigleyC.A., De-BellisA. etal., 1995]. Дигидротестостерон обладает большей афинностью к АР по сравнению с тестостероном, главным образом за счет того, что тестостерон быстро освобождается из своей связи с рецептором. Другие стероиды, такие как андростендион, ДЭАС, эстрадиол, прогестерон, имеют еще меньшее сродство с АР, нежели тестостерон.
Очевидными андроген-зависимыми органами являются мужские гонады и придатки, наружные гениталии, предстательная железа, семенные пузырьки, мышцы, кожа. Однако АР найден и во многих других тканях: гипоталамусе, гипофизе, почке, селезенке, сердце, слюнных железах [WeinbauerG.F., LuetjensC.M., 2010]. В эмбриональный период андрогены определяют дифференцировку наружных гениталий. В период пубертата обеспечивают формирование мужского фенотипа.
В яичках АР экспрессируется в клетках Лейдига, клетках Сертоли, перитубулярных клетках. Экспериментальные исследования показали, что связь андрогенов с АР клеток Сертоли необходима для созревания сперматоцитов [ChangC., ChenY.T., 2004].
Мышцы имеют очень низкую активность 5 альфа-редуктазы, но высокую активность гидроксистероиддегидрогеназы. В скелетных мышцах может осуществляться конверсия андростедиона втестостерон и эстрогены. Основной андроген мышц тестостерон оказывает прямой анаболический эффект на гладкую и поперечно-полосатую мускулатуру с увеличением мышечной массы и гипертрофией волокон. Количество волокон при этом не изменяется. Как следствие действия тестостерона увеличивается синтез РНК и гликогена поперечно-полосатых мышечных волокон. Отсутствие секреции тестостерона может привести к атрофии мышц [WeinbauerG.F., LuetjensC.M., 2010].
Андрогены и эстрогены стимулируют минерализацию костной ткани, увеличивают костную плотность. Метаболизм кости обусловлен взаимодействием тестостерона и эстрадиола. Тестостерон способствует образованию костной ткани. Эстрадиол тормозит как образование, так и резорбцию кости [ValimakiV.V., AlfthanH. еtal., 2004]. Результатом дефицита половых стероидов является остеопороз. В период пубертата увеличение линейного роста кости прямо коррелирует с увеличением концентрации тестостерона. К завершению пубертата происходит закрытие эпифизарных зон роста. При низкой концентрации гормона эти события могут задерживаться.
В период пубертата происходит тестостерон-зависимый рост гортани в длину. Одновременно с этим происходит увеличение длины и массы голосовых связок, приводящее к понижению голосового регистра [ClaassenH., MonigH., 2006]. После завершения периода полового созревания АР теряется, поэтому модификация голоса не может быть достигнута у взрослых мужчин с гипогонадизмом.
В центральной нервной системе также возможна конверсия тестостерона в дигидротестостерон, однако активность ферментов и распределение рецепторов неоднородно в различных зонах мозга. Развитие мозга по мужскому типу детерминировано действием тестостерона. Половая идентификация, поведение, сексуальная ориентация программируются в течение внутриутробного периода развития [ZitzmannM., 2006]. Существует тесная взаимосвязь между уровнем андрогенов и телесной и духовной деятельностью человека. Андрогены влияют на работоспособность, настроение, ощущение уверенности в себе.
Помимо очевидных точек приложения, андрогены влияют на функцию печени, стимулируя синтез белка. Наличие АР в костном мозге обусловливает стимуляцию функции эритропоэза. Влияние андрогенов на жировой обмен характеризуют исследования, подтверждающие снижение уровня ЛПВП при низком уровне тестостерона. Один из механизмов влияния андрогенов на артериальное давление обусловлен стимуляцией выработки эритропоэтина. С другой стороны, прогипертензивный эффект обусловлен влиянием на ренин-ангиотензин-альдостероновую систему [WeinbauerG.F. etal., 2010].
Заключение. Изменение уровня гонадотропинов и половых стероидов на фоне нормально протекающего развития половой системы обусловлено их взаиморегулирующим влиянием друг на друга. Каждый этап развития контролируется комплексом гормональных факторов. Диагностика патологии полового развития основана на комплексной оценке гонадотропинов, половых стероидов в сыворотке крови пациента и их соотношений. Анализируя гормональный профиль детей и подростков, необходимо учитывать физиологические изменения активности половых желез, определяемые онтогенезом и имеющие характерные особенности.
Литература
- Черных В. Б., Курило Л. Ф. Генетический контроль дифференцировки пола у человека // Генетика. 2001. Т. 37, № 10. С. 1317-1329.
- MacLaughlin D. T., Donahoe P. K. Sex determination and differentiation // N Engl J Med. 2004. Vol. 350, № 4. P. 367-378.
- Семичева Т. В., Баканова Т. Д. Дифференциальная диагностика конституциональной задержки пубертата и гипогонадотропного гипогонадизма у мальчиков // Проблемы эндокринологии. 2004. № 3. C. 21-23.
- Ivell R., Hartung S. The molecular basis of cryptorchidism // J. Molecular Human Reproduction. 2003. Vol. 9, № 4. P. 175-181.
- Hughes I. A., Acerini C. L. Factors controlling testis descent // Eur J Endocrinol. 2008. Vol. 159, suppl. 1. S. 75-82. Epub 2008 Jul 22.
- Thomas G. B., McNeilly A. S., Gibson F., Brooks A. N. Effects of pituitary-gonadalsuppression with a gonadotrophin-releasing hormone agonist on fetal gonadotrophin secretion, fetal gonadal development and maternal steroid secretion in the sheep // J Endocrinol. 1994. Vol. 141 (2). P. 317-24.
- Brook C. G., Brown R. S. Handbook of Clinical pediatric Endocrinology. Blackwell Publ., 2008. 265 p.
- Raine J. E., Donaldson M. D., Gregori J. W. et al. Practical Endocrinology and Diabetes in children. Blackwell Publ., 2006. 237 p.
- Suomi A. M., Katharina M. M., Kaleva M., Schmidt I. M. et al. Hormonal Changes in 3-Month-Old Cryptorchid Boys // J of Clinical Endocrinology et Metabolism. 2006. Vol. 91, № 3. P. 953-958.
- Pitteloud N., Hayes F., Boepple P., De Cruz S. The Role of Prior Pubertal Development, Biochemical Markers of Testicular Maturation, and Genetics in Elucidating the Phenotypic Heterogeneity of Idiopathic Hypogonadotropic Hypogonadism // J. Clinical Endocrinology & Metabolism. 2002. Vol. 87, № 1. P. 152-160.
- Zivkovic D., Hadziselimovic F. Development of Sertoli Cells during Mini-Puberty in Normal and Cryptorchid Testes // J Urol. 2009. Vol. 82, № 1. P. 89-91.
- Main K. M., Toppari J., Skakkebek N. E. Gonadal development and reproductive hormones in infant boys // Eur. J. of Endocrinoligy. 2006. Vol. 155. P. 51-57.
- Grinspon R. P., Rey R. A. Anti-Mullerian Hormone and Sertoli Cell Function in Paediatric Male Hypogonadism // Horm. Research in Pediatrics. 2010. Vol. 73 (2). P. 81-92.
- Bougnères P., François M., Pantalone L. et al. Effects of an Early Postnatal Treatment of Hypogonadotropic Hypogonadism with a Continuous Subcutaneous Infusion of Recombinant Follicle-Stimulating Hormone and Luteinizing Hormone // Journal of Clinical Endocrinology et Metabolism. 2004. Vol. 93, № 6. P. 2202-2205.
- Kelly J. A., Vankrieken L. Binding Globulin and the Assessment of Androgen Status / Eur. Eng. Diagnostic Products Corporation, 1999.
- Дедов И. И., Калинченко С. Ю. Возрастной андрогенный дефицит у мужчин. М. : Практическая медицина, 2006. 239 с.
- Дедов И. И., Семичева Т. В., Петеркова В. А. Половое развитие детей: норма и патология. М. : Медицина, 2002. 180 с.
- Подтетнев А. Д., Братчикова Т. В., Орлов Е. Н. Стероидные гормоны и их роль в течение беременности и родов. М., 2000. 222 с.
- Labrie F. Adrenal androgens and intracrinology // Semin Reprod med. 2004. Vol. 22, № 4. P. 299-309.
- Assessment of Circulating Sex Steroid Levels inPrepubertal and Pubertal Boys and Girls by a NovelUltrasensitive Gas Chromatography-Tandem MassSpectrometry Method / F. Courant, L. Aksglaede, J. Antignac [et al.] // J Clin Endocrinol Metab. 2010. Vol. 95, № 1. P. 82-92.
- Talbot N. B., Butler A. M., Berman R. A., Rodriguez P. M., MacLachan E. A. 1943 Excretion of 17-ketosteroids by normal and by abnormal children // Am J Dis Child. 1943. Vol. 65. P. 364-375.
- Albright F., Smith P. H., Fraser R. A syndrome characterized by primary ovarian insufficiency and decreased stature. Report of 11 cases with digression on hormonal control of axillary and pubic hair // Am J Med Sci. 1942. V.204. P. 625-648.
- Kelnar C. J. H. , Brook C. G. D. A mixed longitudinal study of adrenal steroid excretion in childhood and a mechanism of adrenarche // Clin Endocrinol (Oxf). 1983. Vol.19. P. 117-129.
- Wierman M. E., Beardsworth D. E., Crawford J. D., Crigler J. F. et al. Adrenarche and skeletal maturation during luteinizing hormone releasing hormone analogue suppression of gonadarche // J Clin Invest. 1986. Vol. 77. P. 121-126.
- Nakamura Y., Gang H. X., Suzuki T., Sasano H. Adrenal changes associated with adrenarche // Rev Endocr Metab Disord. 2009. Vol. 10. P. 19-26.
- Zemel B. S., Katz S. H. The contribution of adrenal and gonadal androgens to the growth in height of adolescent males // Am J Phys Anthropol. 1986. Vol. 71. P. 459-466.
- Largo R. H. 1993 Catch-up growth during adolescence // Horm Res. 1993. Vol. 39. P. 41-48.
- Ong K. K., Potau N., Petry C. J., Jones R. et al. Opposing influences of prenatal and postnatal weight gain on adrenarche in normal boys and girls // Journal of Clinical Endocrinology et Biochemistry. 2004. Vol. 89. P. 2647-2651.
- Lourdes I. N., Joan D. N., Potau N., SaengerPPremature Adrenarche—Normal Variant or Forerunnerof Adult Disease? // Endocrine Reviews. 2000. Vol. 21, № 6. P. 671-696.
- Sorensen K., Andersson A. M., Skakkebek N. E., Juul A. Serum Sex Hormone-Binding Globulin Levels in Healthy Children and Girls with Precocious Puberty before and during Gonadotropin-Releasing Hormone Agonist Treatment // Journal of Clinical Endocrinology et Metabolism. 2007. Vol. 92, № 8. P. 3189-3196.
- Wilson J. D. Role of dihydrotestosterone in androgen action // Рrostate Suppl. 1996. Vol. 6. P. 88-92.
- Zhu Y. S., Imperato-McGinley J. L., Ann N.Y. 5 alpha-reductase isozymes and androgen actions in the prostate // Acad Sci. 2009. Vol. 1155. P. 43-56.
- Quigley C. A., De-Bellis A. et al. Androgen receptor defects: historial, clinical and molecular perspectives // Endocr. Rev. 1995. № 16. P. 271-321.
- Nieschlag E., Behre HM, Nieschlag S. Andrology: Male repropdactive health and dysfunction. 2010. P. 49-61.
- Chang C., Chen Y. T., Yeh S. D. et al. Infertility with defective spermatogenesis and hypotestosteronemia in male mice lacking the androgen receptor in Sertoli cells // Proc Natl Acad Sci USA. 2004. Vol. 101. P. 6876-81.
- Valimaki V. V., Alfthan H., Ivaska K. K. et al. Serum estradiol, testosterone, and sex hormone-binding globulin as regulators of peak bone mass and bone turnover rate in young Finnish men // J Clin Endocrinol Metab. 2004. Vol. 89. P. 3785-3789.
- Claassen H., Monig H., Sel S., Werner J. A., Paulsen F. Androgen receptors and gender-specifi c distribution of alkaline phosphatase in human thyroid cartilage // Histochem Cell. Biol. 2006. Vol. 126. P. 381-388.
- Zitzmann M. Testosterone and the brain // Aging Male. 2006. Vol. 9. P. 195-199.
- Меньшиков В. В. Стандартизациявклиническойлабораторной медицине. Организационные и метродологические аспекты / под ред. В. В. Меньшикова. М., 2005. 251 с.
- Courant F., Aksglaede L., Antignac J. P., Monteau F., Sorensen K. et al. Assessment of Circulating Sex Steroid Levels in Prepubertal and Pubertal Boys and Girls by a Novel Ultrasensitive Gas Chromatography-Tandem Mass Spectrometry Method // J Clin Endocrinol Metab. 2010. Vol. 95 (1). P. 82-92.
- Карпищенко А. И. Медицинские лабораторные технологии. СПб. : Интермедика, 2002. Т. 2. 470 с.
- Урусова М. Е. Сравнительный анализ определения концентрации ряда аналитов в сыворотке крови с использованием коммерчески доступных наборов реагентов четырех фирм-производителей (Алкор-Био, Roche, DiagnosticProductsCorporation, BayerCorporation) // Клиническая лабораторная диагностика. 2002. № 4. С. 16-35.
- Chada M., Prusa R., Bronsky J. Inhibin B, Follicle Stimulating Hormone, Luteinizing Hormone and Testosterone during Childhood and Puberty in Males: Changes in Serum Concentrations in Relation to Age and Stage of Puberty // Physiol. Res. 2003. Vol. 52. P. 45-51.
- Lahlou N., Fennoy I., Carel J. C. et al. Inhibin B and Anti-Mullerian Hormone, But Not Testosterone Levels, Are Normal in Infants with Nonmosaic Klinefelter Syndrome // The Journal of Clinical Endocrinology & Metabolism. 2004. Vol. 89 (4). P. 1864-1868.