Теоретические и прикладные аспекты исследования функциональной активности политенных хромосом под влиянием разных факторов
ID: 2015-06-1276-A-5303
Оригинальная статья (свободная структура)
ГБОУ ВПО Саратовский ГМУ им. В.И. Разумовского Минздрава России
Резюме
нет
Ключевые слова
хромосомы
Статья
Введение. Изменение функциональной активности хромосом (ПХ) относится, как правило, к ненаследственной, обратимой или эпигенетической изменчивости, часто возникающей в результате изменения экспрессии гена или группы генов (без изменения структуры генетического материала), например, вследствие метилировании ДНК и (или) компактизации хроматина [112]. Изменение активности хромосом в интерфазном периоде лежит в основе изменения функционирования генома в целом и может приводить к ряду патологических состояний клеток, например, к малигнизации соматических клеток млекопитающих. Так, примерно в 20% случаев наследуемого рака обнаруживается взаимосвязь между уровнем метилирования определенных локусов и полиморфизмом генов, связанных с риском онкологического заболевания [123]. Кроме того, изменение ФА генов, инициируя целый каскад биологических реакций как на клеточном, так и органном уровнях, может находиться в прямой взаимосвязи со структурными повреждениями ДНК и участками хромосом, являясь не только их следствием (эффект положения гена), но и одной из причин их возникновения (например, за счет активации т.н. горячих точек хромосом). Например, появление клеточных клонов с новыми хромосомными перестройками способствует опухолевой прогрессии [88, 89, 132, 133].
С другой стороны, изменение плоидности ядер в клетках опухоли и возникновение в них политенных хромосом (ПХ) может рассматриваться одним из признаков их злокачественного перерождения, наряду с гетерогенностью опухолевых клеток по числу хромосом, структурным перестройкам и псевдоплоидии [24, 87, 132, 134], в связи с чем гигантские хромосомы становятся объектом исследования онкологов. Кроме того, характер активности работающих на определенном этапе онтогенеза генов может иметь непосредственное значение для прогнозирования развития заболевания и выбора оптимальной стратегии лечения.
Между тем, до сих пор не установлены те изменения ФА интерфазных хромосом, которые могут приводить к необратимой патологии клеток, например, малигнизации с их дальнейшей опухолевой прогрессией, в связи с чем необходимы определения четких параметров ФА интерфазных хромосом, накопление и анализ данных об изменении значений таких параметров под действием разных факторов, воздействующих на хромосомный аппарат соматических клеток эукариотических организмов.
Наиболее пригодным модельным объектом для анализа ФА интерфазных хромосом эукариот служат ПХ или гигантские хромосомы, постоянно находящиеся в интерфазном периоде [46]. У млекопитающих такие хромосомы обнаружены в трофобластах и гигантских раковых клетках [33]. Наиболее удобным модельным объектом служат ПХ ряда специализированных тканей Diptera – дрозофил, кулицид, мошек, хирономид [47]. Размер и структура ПХ позволяют проводить их идентификацию в кариотипе по рисунку дисковой исчерченности, выявлять инверсии, делеции и дупликации целых участков, проводить наблюдения за изменением ФА отдельных генов в норме и под воздействием разных факторов. Максимальную степень политении достигают хромосомы из клеток слюнных желез личинок Сhronomus (Сhironomidae, Diptera) IV возраста [73, 78, 79].
Цель настоящей работы: определить показатели ФА интерфазных хромосом соматических клеток на примере ПХ личинок двукрылых насекомых (Diptera) как проявление эпигенетической изменчивости, представить и проанализировать данные по изменению показателей такой активности как под действием факторов с уже известным действием на генетический аппарат или матричный синтез на других группах эукариотических организмов, так и еще с неустановленным.
Результаты и их обсуждение. Строение политенных хромосом и параметры их функциональной активности. Гигантские хромосомы у хирономид, впервые описанные Бальбиани в 1881 году, представляют собой длинные канатообразные и поперечно исчерченные образования, которые в зависимости от стадии развития личинки могут иметь различный внешний вид: ленты, меандра, или цилиндра [33, 35]. Максимальная степень политении достигается на конечных этапах дифференцировки, незадолго до гибели органа. У личинок IV возраста, обычно используемых в хромосомном анализе, ПХ имеют вид цилиндра. Большие размеры ПХ обусловлены соматической конъюгацией гомологов, а также тем, что каждая ПХ в результате последовательных актов репликации конъюгирующей диплоидной пары оказывается многонитчатой; при этом, реплики не разделяются, а остаются прикрепленными друг к другу по всей длине; а также их перманентным состоянием в интерфазе [33, 35, 47]. Основным структурным элементом ПХ служит волокно ДНП (хроматида), диаметр которого колеблется от 11 до 25 мкм [135]. ПХ представлены длинными лентами с отчетливыми полосами (рис. 1), которые образованы поперечными дисками, состоящими из продольно соединенных между собой хромомер. Хромомер - отрезок компактизованной ДНК, выступающий как общебиологический механизм обеспечения устойчивости организма к неблагоприятным условиям.

Рис. 1. Структура политенной хромосомы III (EF) Chironomus riparius в контроле.Стрелкой обозначена центромера.
Рисунок дисков ПХ уникален и видоспецифичен. Количество дисков таких хромосом у хирономид варьирует от 1.5 до 2.5 тысяч [35, 113, 122]. Диски пассивно создаются как фрагменты неактивной в транскрипции ДНК между двумя активными междисками: они образованы участками хроматина с наиболее плотной конденсацией ДНП [84], содержат гены и фрагменты ДНК, нетранскрибируемые в данной ткани в данный момент; материал этих генов находится в компактном состоянии. Диски окрашиваются более интенсивно по сравнению с междисками и выявляются как более плотные участки, возникающие вследствие локального увеличения плотности упаковки индивидуальных хроматиновых нитей в хромомерах. Размеры и морфология дисков, расстояния между соседними дисками строго индивидуальны для каждого даже небольшого фрагмента хромосом. В результате отдельные группы дисков могут служить маркерами не только районов, но и целых хромосом. По-видимому, диски служат информативными единицами хромосом и могут рассматриваться как места расположения основной массы генов и как единицы транскрипции и репликации [47].
Структура дисков может изменяться при резкой перестройке метаболизма клеток слюнных желез. При этом диски, сливаясь, формируют блоки конденсированного хроматина, в результате хромосомы укорачиваются в два-три раза, что сопровождается инактивацией синтеза РНК [36]. Ряд химических агентов может приводить к грануляции дисков, увеличению участков междисковых пространств, разворачиванию хромосом в ленты-меандры; все это в итоге ведет к дезорганизации структуры и функционирования интерфазных хромосом [47, 73. 101, 102], поэтому длина хромосомы и показатели ее компактности используются при оценке ФА ПХ.
Междиски - транскрипционно активные районы ПХ, ответственные за функции основного метаболизма клеток (содержат работающие гены «домашнего хозяйства») [34]. Доказано [38], что междисковые области активно транскрибируются и, по-видимому, контролируют основной метаболизм в клетках.
Для ПХ хирономид характерно наличие активных районов (рис. 2) – ядрышковых организаторов (ЯО), пуфов и колец Бальбиани (КБ). Число и локализация ЯО и КБ в ПХ Chironomini видоспецифичны и используются как видовой признак. В ядре - одно или несколько ЯО. В ЯО находятся серии генов, кодирующих рРНК; кроме того, здесь происходит образование субъединиц рибосом. Фенотипическим проявлением активности ЯО являются ядрышко и ядрышкоподобные структуры, которые доступны для количественного и качественного анализов. Экспрессия рибосомного локуса служит удобной естественной моделью, позволяющей проводить количественную оценку проявления в фенотипе как генетических его особенностей, так и реакции на условия среды обитания. При изучении ЯО возможен анализ ФА четко локализованных генов, ответственных за синтез предшественников рРНК (38S, 23S, 30S) [34].
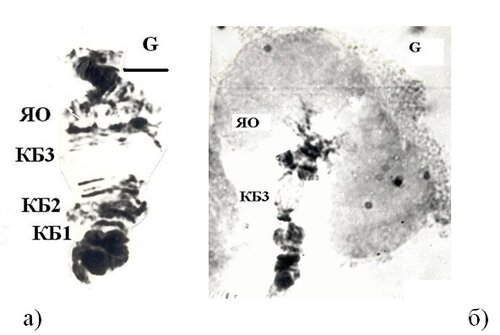
Рис. 2. Структура политенной хромосомы IV (плечо G) Chironomus riparius в контроле (а) и индукция ЯО и КБ3 хромосомы IV (б).
В клетках с ПХ ядрышки выглядят крупными округлыми образованиями, отличными по консистенции от остальной нуклеоплазмы. Размер ядрышка уменьшается в конце последней личиночной стадии. ЯО типа Chironomus картируется в определенном районе; в полностью неактивном состоянии зона ЯО имеет четкий дисковый рисунок.
В результате активации генов, расположенных в дисках, материал ПХ разрыхляется, декомпактизируется и морфологически увеличивается в размерах, образуется вздутие – пуф. Выделяют несколько типов пуфов – КБ, ДНК–пуфы и обычные пуфы. Наиболее крупные пуфы - Кольца Бальбиани – с особым расположением хроматид в активном районе характерны для таких двукрылых насекомых, как хирономиды, симулииды, кулициды, реже встречаются у дрозофил. Пуфы, в которых происходит амплификация ДНК и затем синтезируется РНК, встречаются, в основном, у сциарид (ДНК-пуфы); они появляются в конце личиночного развития, участвуют в подготовке клеток к гистолизу и кодируют полипептиды секрета. Обычные пуфы описаны для всех видов животных и растений, имеющих ПХ классического типа.
Цикл развития пуфа состоит из следующих этапов: на первом - один диск или группа компактных сильно окрашивающихся дисков разрыхляется, степень окрашиваемости уменьшается (диски становятся «серыми» под фазовоконтрастным микроскопом); на втором - одновременно с увеличивающимся в размерах вздутием исчезают очертания дисков. Степень декомпактизации материала в развитом пуфе максимальна, пуф теряет окрашиваемость, становится «белым» при окраске или под фазовым контрастом [33]. Во время регрессии пуфа следуют те же этапы, но в обратном порядке: сначала уменьшается вздутие, затем материал полностью компактизируется в диск.
Пуфы служат показателями дифференциальной генной активности; в них происходит активный синтез РНК [47, 114]. В пуфе, в отличие от диска, отсутствует плотная упаковка фибрилл ДНП [47]. Спектр пуфов в норме строго характерен для каждой ткани на каждом этапе онтогенеза [1, 114]. Нарушение общей сбалансированности работы пуфов может приводить к возникновению новых, неспецифических для данного организма, пуфов denovo [47]. Факторами, влияющими на пуфинг в целом, выступают, в частности, гормон экдистерон и тепловой шок. В т.н. пуфах теплового шока у Chironomidae образуются гигантские гранулы РНП. Пуф-индуцирующая активность была продемонстрирована для аминокислот, бензамида, диэтилового спирта, хлороформа, бромдезоксиуридина, внутриклеточных инфекций, гемолимфы имаго, гиббереллиновой кислоты, дигитонина, колхицина, ментола, окситетрациклина, пактамицина, пуромицина, сахаров (галактозы, сахарозы, трегалозы), спиртов, различных инкубационных сред, циклогексимида.
КБ представляют собой вздутия ПХ, затрагивающие до 10-20 дисков [39, 47]. Отличием КБ как гигантского пуфа от типичных пуфов является большая конденсация хромонем, имеющих вид крупных петель и выступающих за пределы ПХ [47]. ДНК КБ состоит из многочисленных повторяющихся элементов и кодирует гигантские по размерам молекулы РНК для синтеза структурных белков секрета слюнных желез, необходимого для построения личиночного домика, поэтому КБ находятся в активном состоянии в ПХ клеток слюнных желез на всех личиночных стадиях развития [33, 47]. Транскрипция в КБ происходит только на личиночной стадии развития и только в слюнных железах, позже – инактивация. В процессе образования КБ узколокальный участок хромосомы декомпактизуется, превращаясь в небольшое вздутие, которое быстро растет в диаметре (рис. 2). По обе стороны от этого вздутия хромосома расщепляется на хроматиды, образующие петли, большое число которых образует муфтообразную структуру, как бы нанизанную на хромосому.
Помимо функционально активных районов ПХ реакции ядерного генома могут проявляться и морфологическими изменениями неактивных участков – гетерохроматиновых районов. Гетерохроматиновые сегменты представляют собой материал, дополнительный к эухроматину. Гетерохроматин (ГХ) всегда конденсирован, генетически неактивен, в большинстве случаев не транскрибируется, реплицируется в конце синтетического периода интерфазы, в его состав входят часто повторяющиеся последовательности нуклеотидов, не кодирующие белки. ГХ интенсивно окрашивается, потеря его участков может не отражаться на жизнедеятельности клетки. Количество ГХ, а, соответственно, величина гетерохроматиновых сегментов может варьировать [91]. Такое явление, называемое полиморфизмом гетерохроматиновых сегментов, носит, по-видимому, всеобщий характер и дает возможность иметь маркеры для определения родительской принадлежности гомологичных хромосом.
Известна неспецифичная реакция ГХ на разные факторы среды (высота, температура и соленость водоема), в основе которой, по-видимому, лежат механизмы регуляции конденсации–деконденсации ГХ через ядерную мембрану [34]. Так, повышенное содержание ГХ добавочной В-хромосомы характерно для Сhironomusbehningi, обитающей в солоноватых водоемах [9]. Влияние высоты над уровнем моря (свыше 2000 м) на увеличение количества ГХ в хромосомах было отмечено у первичнобескрылых насекомых [34]. Кроме того, присутствие в хромосомах ГХ обеспечивает высокую частоту хромосомных перестроек, а изменение его содержания у разных видов, по мнению Прокофьевой-Бельговской [85], имеет приспособительное значение и обеспечивает быструю адаптацию к изменяющися условиям среды быстрее, чем мутационный процесс. Изменение содержания ГХ в геноме хирономид, вероятно, происходит путем дупликаций генетического материала, которым придается большое значение в эволюции [126]. Наклонность ГХ к слипанию по принципу подобного с подобным [34] при низких температурах способствует объединению хромосом в хромоцентр, реципрокному обмену хромосомных плеч и возникновению новых цитокомплексов [108]. За счет гетерохроматиновых участков осуществляются эктопические контакты между негомологичными хромосомами [33, 34, 63, 32].
Для большинства Chironomini характерен хромосомный полиморфизм инверсионного типа [12, 50, 60, 61, 63, 71, 106, 126]. Инверсионные разломы в хромосомах в основном приходятся на строго определенные диски или междиски – т.н. «горячие точки» в геноме, представляющие собой участки интеркалярного гетерохроматина, не полностью реплицирующиеся в ходе политенизации [38, 61]. Благодаря принципу эволюционной уникальности хромосомных перестроек их используют для реконструкции филогенетических отношений между видами [67, 109].
Таким образом, морфологическими критериями ФА ПХ служат три типа активных районов ПХ – ядрышковый организатор (ЯО), кольца Бальбиани (КБ) и пуфы. Следует учесть, что КБ относятся к специфическим участкам определенных высокоспециализированных тканей, например, слюнных желез двукрылых насекомых, поэтому изменение их активности можно рассматривать как частный случай активности работающих генов двукрылых насекомых на стадии личинки. В таблице 1 представлены морфофункциональные особенности ряда активных участков ПХ, в таблице 2 - метрические показатели и индексы ФА ПХ на примере Ch. plumosus: индекс компактности хромосомы III; коэффициенты ФА ЯО хромосомы IV, КБ1 и КБ2 хромосомы IV; КБ видоспецифичного пуфа плеча В хромосомы I.
При анализе препаратов ПХ целесообразно подбирать однотипные по степени политении хромосом клетки. Шкала степени конденсированности ПХ разработана С.Ю. Деминым [25], согласно которой ПХ могут относиться к пяти классам конденсированности – сильно удлиненными, удлиненными, нормальными (типическими), компактными, сверхкомпактными.
Реакции ядерного генома соматических клеток на воздействие разных факторов среды
Исследование реакции генома эукариотических организмов на воздействие разных факторов среды в настоящее время выделено в отдельное направление - экологическую кариологию, которое, несмотря на большое теоретическое и прикладное значение, пока мало разработано. Именно в рамках этого направления накоплено наибольшее количество данных по изменению состояния ФА интерфазных хромосом под действием разных факторов среды [64- 66, 68-70, 72-79, 81, 94-102, 131].
Накоплены определенные сведения о влиянии на структуру и ФА ПХ хирономид температурного фактора в разные сезоны года [26, 42, 43], а также в условиях вечной мерзлоты Исландии, Гренландии, арктической Аляски, Финляндии, Якутии, севера России [3, 51, 53, 103, 139] и высоты над уровнем моря выше 200 м [34, 44, 80]; других физических и химических факторов [65, 66, 74].
Нами проведена оценка состояния ФА ПХ под действием ксенобиотиков, физических и химических и других факторов по цитогенетическим и генетическим эффектам как специфичным только для данной группы эукариотических организмов (увеличение активности КБ; увеличение активности работы уже имеющихся пуфов; депрессия имеющихся в кариотипе пуфов; инактивация КБ), так и эффектам, которые можно рассматривать универсальными реакциями генома эукариот: снижение или повышение активности ЯО; образование дополнительных ядрышек; появление пуфов denovo; конденсация и укорочение хромосом; деконденсация хромосом; изменение структуры дисков; активация предтеломерных участков; асинапсис мозаичного характера гомологов хромосом; усиление адсорбции красителя; затемнение междисков; меандризация хромосомных участков; разломы хромосом; слияние дисков в блоки, «оплывание» хромосом; эффект «размазывания»; а также возникновение коротких хромосомных аберраций и полиплоидии (табл. 3).
Влияние физических факторов среды на структуру и функциональную
активность ПХ
Влияние температурного фактора. ПХ наименьшей компактности обнаружены у личинок хирономид зимой и ранней весной при низких температурах [25, 41]. При низкой температуре для Chironomus в условиях севера была отмечена эктопическая конъюгация ПХ – центромерные районы их объединялись, образуя хромоцентр, или же хромосомы соединялись друг с другом теломерными районами [51, 53]. Резкое уменьшение числа и размеров пуфов, размеров КБ, уменьшение числа дисков выявлено у Camptochironomus tentans и Ch. plumosus, находящихся в состоянии диапаузы [25, 41, 121, 125]. Искусственное воздействие на личинок хирономид низкими температурами также прекращало работу КБ, создавало картину хромосом, обычно наблюдаемую у молодых куколок, когда ингибирован весь пуффинг, а свою активность сохраняли пуфы, характерные для всего цикла развития [47].
При повышении температуры воды в пределах нормы у личинок Ch. plumosus наблюдалось увеличение числа особей с более конпактными ПХ [25, 41]. ПХ наибольшей компактности обнаружены в конце лета и ранней осенью [26, 40, 42, 43]. Укорочение длины и плохая структурированность ПХ отмечена у хирономид, собранных летом даже в водоемах Полярного Урала, Ямала, Исландии, Гренландии, арктической Аляски, Финляндии [3, 4, 103, 139]. Для ПХ северных видов Chironomus и Camptochironomus характерно сближение дисков, вследствие чего ПХ сильно укорачивались и приобретали крайнюю степень компактности [103]. Значительное укорочение ПХ, конкнсация хроматина и появление пуфов denovo описано у личинок хирономид из Иваньковского водохранилища у Конаковской ГРЭС и из водоема-охладителя Балаковской АЭС при тепловом загрязнении водоема [10, 14, 69, 70].
Сезонные изменения дисков обусловлены изменением степени спирализации ПХ и слиянием групп дисков в плотные, интенсивно окрашивающиеся блоки осенью и разделением их зимой. Диски, сливаясь, образовывали блоки конденсированного хроматина и хромосомы укорачивались в два-три раза. Конденсация хромосомного материала сопровождалась инактивацией синтеза РНК [36]. Локальная конденсация хромосом связана с транскрипцией, поэтому изменение компактности ПХ можно рассматривать как механизм, обеспечивающий гомеостаз клетки при изменении условий обитания личинок [41].
Полученные данные по воздействию на геном Chironomidae температурного шока [31] свидетельствуют в пользу универсальности явления индукции генетической активности на изменение температуры и его адаптивном значении, по крайней мере, для двукрылых насекомых. При стрессовом воздействии перепада температур структурные изменения возникали на всех уровнях упаковки ДНП. Нарушалась упаковка «ленты» в цилиндрическую хромосому: ПХ переходила из цилиндрического состояния в состояние «меандров» или плоской ленты. При воздействии температуры до 36°С отмечены индукция пуфа denovo, увеличение ЯО и КБ1,2, появлялась сегментация в этих локусах, а также грануляция дисков, особенно выраженная в центромерных районах и увеличение междисковых участков ПХ. Такие изменения ПХ были обратимы в пределах 1–3 ч при температуре содержания личинок 18°С. Воздействие температуры свыше 36°С приводило к необратимым последствиям: резкой индукции дисковой грануляции, увеличению междисков и раскручиванию ПХ, а также индукции экспрессии генов белков теплового шока [31].
Влияние фактора высоты над уровнем моря Влияние высоты над уровнем моря (свыше 2000 м), в отношении гидробионтных личинок гетеротопных двукрылых насекомых, скорее всего, сводится к воздействию на них преимущественно таких факторов, как резкие суточные колебания температуры, пониженное содержание кислорода и ультрафиолетовое излучение, характерные для таких высот. Так, хирономидам, обитающим в условиях вечной мерзлоты с пониженной температурой, свойственно повышенное количество прицентромерного ГХ [ ] (Кикнадзе с соавт., 1998). Влияние низкой температуры на структуру хромосом северных Сhironomus выражено в разрывах хромосом, образованию транслокаций, появлению множественных В-хромосом, состоящих из ГХ [52, 72, 108]. Нами у личинок Chironomus горных и высокогорных популяций отмечена фиксация в кариотипе тех гомозиготных хромосомных последовательностей, которые редко встречались у личинок этих видов из равнинных популяций даже в гетерозиготном состоянии [44, 80].
Влияние фактора глубины. Укороченными с высокой степенью конденсации ПХ обладали личинки Ch. plumosus и Stictochironomus pictulus из озера Иссык-Куль, пойманные на глубинах до 61 м [3]. Обитание в глубоководном озере с температурной стратификацией, по-видимому, вызывало приостановку развития (до одной генерации в два года у Ch. plumosus) и изменение метаболизма, что, вероятно, взаимосвязано с конденсацией ПХ.
Механические повреждения клеток. Существуют данные о возникновении крупных перестроек ФА генома в результате повреждения клеточной оболочки в ходе изоляции желез и отдельных клеток [110].
Электромагнитное излучение КВЧ. На Ch. plumosus под действием электромагнитного излучения КВЧ [17] были установлены следующие морфологические изменения ПХ. Отмечалось стимулирующее влияние на функционально активные и предтеломерные участки. Наиболее выраженный биоэффект наблюдался при частоте fc 40.25 ГГц, что проявлялось в изменении компактности ПХ и индукции пуфов denovo. На основе разной активации генома у разных особей одной экспериментальной группы было предположено наличие индивидуальной чувствительности к КВЧ-излучению.
Радиоактивное влияние. Как известно [20, 55, 62, 111], действие радиации оказывает наиболее сильное повреждающее воздействие на молекулы ДНК, вызывая наиболее серьезные изменения в генетическом аппарате клетки. Анализ ФА ПХ под действием факторов, обладающих уже доказанным повреждающим воздействием на ядерный геном эукариот, позволит выделить именное те параметры, на которые следует обращать внимание в первую очередь. В кариотипе хирономид из популяций, подвергшихся радиационному воздействию [3, 5-7, 30, 116], были установлены следующие морфологические изменения ПХ: разрыхленность теломерных районов хромосом, асинапсис мозаичного характера гомологов хромосомы IV (вплоть до полного их расхождения); кроме того, несколько личинок были полиплоидами или обладали характерными микроинверсиями даже в консервативных хромосомных плечах.
В результате наиболее значимыми реакциями ядерного генома, помимо увеличения плоидности (геномная мутация) и микроинверсий (хромосомные аберрации), по-видимому, являются такие изменения ФА ПХ, как конденсация и, соответственно, укорочение хромосом, активация предтеломерных участков, в которых обычно локализованы гены белков теплового шока, и асинапсис мозаичного характера гомологов хромосом, в основе которого также могут лежать мелкие перестройки хромосом, возникшие в результате повреждения молекулы ДНК.
Влияние солености водоема. При увеличении солености ПХ Chironomus valkanovi укорачивались, приобретая рыхлую структуру со сближенными дисками [2, 127, 128]. ПХ хирономид солоновато-водного комплекса из Каспийского моря и гипергалинных водоемов – Chironomus(Halochironomus) sp., Ch. valkanovi, Ch. salinarius, Ch. anchialicus, Baeotendipesnostivaga, B. sp. и др. [68. 129, 130] – в разные сезоны года короткие, с меандрической структурой, вакуолизированные и с распуфленными дисками, что приводило к потере их дискового строения. Для Сhironomusbehningi, обитающей в солоноватых водоемах, также характерно повышенное содержание ГХ добавочной В-хромосомы [9].
Изменение функциональной активности ПХ под действием природных антибиотиков, ксенобиотиков и других химических факторов
Гормональное воздействие. Длительное культивирование слюнных желез в брюшке имаго приводит к резкой перестройке метаболизма клеток, в результате чего также происходит изменение структуры дисков ПХ [37]. Наблюдаются эффекты, сходные с инфекционным воздействием.
Действие природных антибиотиков, лекарственных препаратов и других химических факторов. На Ch. riparius под действием ряда фармацевтических препаратов [11, 73] были установлены общие закономерности – низкие концентрации веществ приводили к активации районов ПХ, а более высокие – к их ингибированию и сблоченности дисков. Такое изменение ФА ПХ, как укорочение за счет слияния соседних дисков, грануляция и сблоченность проявились при воздействии всеми исследованными веществами, что, по-видимому, можно считать общим механизмом ответной реакции интерфазных хромосом эукариот на подобное воздействие.
Действие шести ксенобиотиков (апромицина, тобрамицина, синтанола, 3,5 ксиленола, алкамона, Na-полистерол – сульфатсернистого). Анализируя цитогенетические эффекты шести исследованных нами токсикантов на ПХ Chironomusriparius [73] нами были выявлены следующие закономерности: одни и те же проявления состояния ФА ПХ могут быть вызваны разными токсикантами, что свидетельствует о неспецифической реакции генома в целом; специфическая реакция ПХ может быть связана как с появлением пуфов denovo, так и с определенным спектром показателей состояния генетической активности хромосом при разных концентрациях и экспозициях того или иного токсиканта; наибольшее число проявлений состояния генетической активности ПХ выявлено у тобрамицина, наименьшее у Na-полистерола – сульфатсернистого; такие ксенобиотики, как алкамон, 3,5 ксиленол, апромицин и синтfнол вызывают заметное изменение активности хромосом в первые сутки воздействия; другие ксенобиотики, как Na-полистерол – сульфатсернистый и тобрамицин более резкое изменение ФА ПХ вызывают в последующие 4-6 суток непрерывного воздействия. Снижение активности ЯО, КБ3, пуфов отмечено при воздействии на ПХ 4 ксенобиотиков. Активация КБ1 хромосомы IV и активация теломерных районов хромосом отмечены только при воздействии тобрамицина. В настоящее время показано [48], что КБ, в которые входят тканеспецифические гены, секретируют высокомолекулярную РНК, которая в составе РНП-гранул попадает в цитоплазму и служит матрицей для синтеза секреторных белков. Активность КБ в клетках слюнных желез контролируется несколькими регуляторными системами. С одной стороны, это автономный механизм, функционирующий по принципу обратной связи, в зависимости от степени наполнения протоков слюнных желез секретом, с другой – работа данного механизма зависит от стадии развития личинок [47].
Такие изменения ФА ПХ, как их укорочение за счет слияния соседних дисков, грануляция и сблоченность проявились при воздействии всех ксенобиотиков. Укорочение ПХ, по-видимому, является общим механизмом ответной реакции интерфазных хромосом на воздействие окружающей среды. Увеличение активности ЯО, КБ3, пуфов было вызвано четырьмя из шести изученных ксенобиотиков. Усиление активности ЯО может быть связано с повышением устойчивости клетки к внешним воздействиям и рассматриваться как адаптивная реакция, тогда как снижение ФА этого локуса под влиянием неблагоприятных факторов может внести свои коррективы в гомеостаз популяции [45].
При стрессе очень быстро изменяется экспрессия генов, что приводит к образованию стрессовых белков. Биохимические функции стрессовых белков неясны, но с ними связывают становление устойчивости клетки к стрессирующему фактору. Индукция активного района, однако, еще не означает синтеза РНК в нем, т.е. деконденсация хроматина и синтез РНК являются, хотя и взаимосвязанными, но самостоятельными процессами. Так, при изучении эффектов действия актиномицина D на транскрипцию в ПХ [19] было обнаружено неожиданное свойство сохранения или даже индукции пуфов при полном ингибировании синтеза ДНК высокими дозами антибиотика (10–25 мкг/мл). Синтез стрессовых белков сопряжен с изменениями степени конденсированности ПХ [31]. Возможно морфологическим свидетельством экспрессии генов, отвечающих за синтез подобного рода белков, является образование пуфов denovo в ПХ, распуфливание предтеломерных районов под воздействием ряда ксенобиотиков.
Индукция пуфов denovo была вызвана двумя ксенобиотиками – тобрамицином и апромицином. Известно, что при стрессе очень быстро изменяется экспрессия генов, приводя к образованию стрессовых белков. Биохимические функции стрессовых белков неясны, но с ними связывают становление устойчивости клетки к стрессирующему фактору. Индукция активного района, однако, еще не означает синтеза РНК в нем, т.е. деконденсация хроматина и синтез РНК являются, хотя и взаимосвязанными, но самостоятельными процессами. Так при изучении эффектов действия актиномицина D на транскрипцию в ПХ [19] было обнаружено неожиданное его свойство сохранять или даже индуцировать пуфы при полном ингибировании синтеза ДНК высокими дозами антибиотика (10-25 мкг/мл). Но синтез стрессовых белков всегда сопряжен с изменениями степени конденсированности ПХ, поэтому возможно, что образование пуфов denovo в гигантских хромосомах, распуфливание предтеломерных районов под воздействием тобрамицина, апромицина и является морфологическим свидетельством экспрессии генов, отвечающих за синтез подобного рода белков.
Влияние атропина и пилокарпина. Известно [93], что у личинок хирономид активен фермент – холинэстераза; благодаря холинергической нейромедиации ПХ клеток слюнных желёз как секреторного органа, являются наиболее чувствительными к воздействию таких препаратов. как атропин и пилокартин. Под действием пилокарпина секрет из слюнных желез личинок Ch. riparius и Camptochironomustentans полностью выводился [113]; во время же восстановления секреции увеличивалась ФА пуфов и колец Бальбиани (КБ) хромосомы IV [19]. Введение атропина в организм личинок, наоборот, сопровождалось уменьшением секреции. Неодинаковая реакция активных районов и компактности ПХ Ch. plumosus [94, 101, 102] на воздействие атропина связана с их различной ролью в поддержании гомеостаза, как клетки, так и организма в целом. Ответная реакция участков «специфических функций» на действие атропина неоднозначна, изменения ФА КБВ, КБ1G и КБ2G несогласованны между собой, что свидетельствует о разбалансировке секреторной функции желез, особенно в период между острым и подострым воздействием, когда происходит «настройка» внутриклеточных репаративных механизмов. ФА ЯОG более чувствительна к изменению среды по сравнению с компактностью. При длительном воздействии репрессия работы КБВ, КБ1G и КБ2G сопровождалась снижением секреции слюнных желез, что согласуется с действием атропина на слюнную железу. Атропин вызывал появление пуфов denovo, не отмеченных в контроле и возникающих произвольно у единичных особей. Изменения ФА по индексу видоспецифичного пуфа плеча в в целом не коррелируют с работой других активных участков ПХ.
При воздействии пилокарпина реакция ПХ Ch. plumosus [94, 101, 102] по индексу компактности неадекватно отражает изменения ТА. Угнетение ФА ЯО при действии высоких концентраций препарата в период подострого воздействия служит критерием подострого токсического действия пилокарпина. Отсутствует чёткая корреляция в работе КБВ и КБG между собой. Пилокарпин изменяет дифференциальную генную активность, индуцируя появление пуфов denovo, не отмеченных в контроле. При всех исследованных концентрациях и экспозициях пуфинговая активность под действием пилокарпина возникает у единичных особей и изменяется произвольно.
Изменение функциональной активности ПХ под действием
биологических и генетических факторов
Инфекция микроорганизмами сопровождалась резкой перестройкой метаболизма клеток слюнных желез Chironomidae, в результате чего меняется структура дисков. Сливаясь, диски формируют блоки конденсированного хроматина, вследствие этого ПХ укорачивались в 2–3 раза; конденсация ПХ сопровождается инактивацией синтеза РНК [37].
Влияние генетического фактора – инбридинга. Как известно [22], при инбридинге родители, являясь родственниками, имеют много одинаковых аллелей, в результате чего гомозиготность увеличивается с каждым поколением. Инбридинг приводит к повышению постоянства фенотипических признаков в потомстве и генетически сходных или идентичных особей (инбредные линии). При близкородственном скрещивании может возникать депрессия: уменьшение размеров животных, возникновение аномалий и уродств, что объясняется гомозиготностью по вредным рецессивным аллелям. На ПХ [57, 58] инбридинг приводил к следующим изменениям – увеличению активности работы уже имеющихся пуфов; депрессии имеющихся в кариотипе пуфов; снижению ФА ЯО; появлению пуфов denovo.
Анализ цитогенетических и генетических эффектов
Анализируя около 40 приведенных в таблице 3 фактов воздействия на ПХ, можно отметить следующее. Наиболее часто повторяющимися цитогенетическими эффектами являются: появление пуфов denovo – 45%, снижение ФА ЯО – 32, конденсация и укорочение хромосом – 23, увеличение активности КБ –19, увеличение активности пуфов –19, депрессия имеющихся в кариотипе пуфов –13. Наиболее редкими показателями изменения ФА ПХ следует считать увеличение объема ядрышка, деконденсация хромосом, активация предтеломерных участков и образование дополнительных ядрышек. Такая ответная реакция генома, как появление пуфов denovo, специфична. Она связана с включением в работу ряда молчащих генов и синтезом белков, необходимых организму, по-видимому, для поддержания гомеостаза в критических ситуациях. Образование пуфов denovo в половине проанализированных фактов воздействия отражает объективное повышение ФА генома в целом.
Анализ работы ЯО обнаружил наиболее частое снижение его активности под действием ксенобиотиков (32%). Конденсация и укорочение хромосом, как ответная реакция на воздействие, проявилась чаще (23%), чем деконденсация ПХ. Анализ изменения структуры и ФА ПХ под влиянием разных ксенобиотиков и других факторов среды, на первый взгляд, может показать неспецифическую реакцию генома, за исключением образования пуфов denovo. Однако анализ спектра значений показателей ФА ПХ внутри разных групп исследованных веществ позволил выявить сочетания цитогенетических эффектов. Такие сочетания могут свидетельствовать не только о единстве механизмов, вызвавших цитогенетические эффекты, но и определить специфичность воздействия на геном тех или иных факторов.
Как было показано нами (табл. 3), наиболее часто встречаются такие сочетания, как появление пуфов denоvo на фоне увеличения активности уже имеющихся пуфов, – 19%, появление пуфов denovo на фоне снижения ФА ЯО – 13, а также снижение ФА ЯО на фоне общей конденсации хромосом и их укорочении – 1. Анализируя выявленные сочетания цитогенетических эффектов, можно полагать, что механизмы активации уже имеющихся пуфов в геноме вида с появлением пуфов denovo функционально связаны между собой. Функциональную связанность можно отметить и в отношении снижения ФА ЯО и конденсации хромосом.
Отсутствие взаимосвязи показывают такие эффекты, как активация ранее молчащих генов (появление пуфов denovo) и активация работы ЯО и КБ, а также общая конденсация – деконденсация ПХ и ФА ЯО. Так, увеличение ФА КБ в четырех из шести фактов проявления отмечено отдельно от других эффектов и только в двух случаях – на фоне снижения активности работы ЯО. Установлено (Кикнадзе, 1985), что КБ, в которые входят тканеспецифические гены, секретируют высокомолекулярную РНК, которая в составе РНП-гранул попадает в цитоплазму и служит матрицей для синтеза секреторных белков. ФА КБ в клетках слюнных желез контролируется несколькими регуляторными системами. С одной стороны, такой регуляторный механизм – автономный, функционирующий по принципу обратной связи, в зависимости от степени наполнения протоков слюнных желез секретом, с другой, – работа данного механизма зависит от стадии развития личинок [47].
При детальном исследовании атропина и пилокарпина на ФА ПХ Ch. plumosus [74, 94, 101, 102] было установлено, что наиболее чувствительны к действию лекарственных препаратов индекс компактности ПХ и индексы ФА ЯО и КБ1 хромосомы IV. Выявлено, что значения показателя «общих функций» (индекса ФА ЯО) меняются адекватно воздействию холинотропных препаратов invivo в отличие от показателей «специфических функций» (индексов ФА КБ и видоспецифичного пуфа), характер изменений которых несогласован. Установлен определенный баланс между активно работающими и неактивными КБ и пуфами denovo, что характеризует работу генома, как сбалансированную систему с компенсаторными механизмами по принципу обратной связи. Определен период наиболее значительного воздействия лекарственных препаратов invivo на генетический аппарат, во время которого происходит нарушение баланса в работе активных участков, - период острого воздействия (до 24 ч.). Выявлено, что цитогенетические эффекты зависят от концентрации действующего препарата invivo: при возрастании концентрации атропина ФА хромосом снижается, а при пилокарпине – повышается. Исключением является угнетение ФА ЯО в подострый период действия пилокарпина (48-72 ч.), что свидетельствует о его токсичности. На субклеточном уровне токсичность препарата проявляется также в увеличении ФА ЯО в период острого воздействия и длительном периоде восстановления активности участков «специфических функций» до состояния контроля. Обнаружено, что с увеличением размера тела личинок отмечено повышение ФА ЯО и пуфа при действии атропина, и ФА КБВ и КБ1 при действии пилокарпина.
Установлено, что при комбинированном влиянии холинотропных препаратов, воздействуя на личинку сначала невысокой концентрацией (1/15 LC50) м-холиноблокатора – атропина в течение 24 ч., затем – м-холиномиметика – пилокарпина (1/15 LC50) наибольший стимулирующий эффект большинства ФА участков ПХ наблюдался только через 48 часов. В смеси препаратов при всех экспозициях ФА большинства участков ПХ была угнетена и деблокада пилокарпином не происходила. При наиболее токсичной концентрации пилокарпина (1/5 LС50), приводящей к образованию максимального количества пуфов denovo, все изменения ФА обратимы, что подтверждает модификационный характер изменчивости морфологии ПХ. Выявлена прямая зависимость между изменениями ФА участков «специфических функций» ПХ и долей гетерозиготных инверсий в кариотипе - при действии атропина с увеличением доли гетерозиготных инверсий в кариотипе повышается ФА двух КБ (КБВ и КБ1G), а при действии пилокарпина - всех трех КБ, показывая устранение разбалансировки в работе активных участков при действии препарата. Присутствие гетерозиготных инверсий в кариотипе показывает преимущество гетерозиготных особей, подтверждая адаптивную роль таких хромосомных перестроек. Для оптимизации метода цитогенетической оценки лекарственных препаратов предлагается использовать комплекс морфометрических показателей ФА, как участков «общих», так и «специфических» функций. Ведущим показателем служит ФА ЯО, работающего наиболее стабильно, в 97–99% клеток, и являющегося одним из показателей адаптации при поддержании клеточного гомеостаза.
Как видно (табл.3) к факторам, способным изменить ФА ПХ и привести к изменению функции участков генома клетки, относятся как химические, физические, так и генетические, биологические, т.е. подавляющее большинство факторов окружающей среды, которым эукариотические организмы подвергаются каждодневно. Перечисленные морфо-функциональные особенности ПХ, возникающие под действием таких факторов, наряду с более крупными перестройками – геномными мутациями, хромосомными аберрациями и появлением в кариотипе добавочных хромосом, по-видимому, представляют собой устойчивые адаптации на клеточном уровне хирономид и эукариотических организмов в целом. Специфичность реакции генома на факторы среды можно установить на основе анализа спектра показателей ФА ПХ, что отражает наличие в геноме тонких механизмов связи функциональных участков между собой. Исследование по установлению корреляций в работе разных участков ядерного генома возможно только на основе комплексного подхода с привлечением биохимических и молекулярно-генетических методов анализа генной активности и мутаций, возникающих на разных уровнях организации наследственного материала – генном, хромосомном и геномном.
К наиболее значимым морфологическим изменениям ФА ПХ, которым следует уделять особое внимание в связи с возможным последующим патологическим состоянием при исследовании интерфазных хромосом соматических эукариотических клеток, подвергшихся определенному воздействию, относят: компактизацию хроматина, активацию ЯО, активацию предтеломерных участков хромосом, а также возникновению микроаберраций, морфологически проявляющихся асинопсисом гомологов.
Литература
- Агапова О. А., Кикнадзе И. И. Пуффинг и специфическая функция слюнных желез Chironomus thummi. III. Ультраструктура клеток при личиночной линьке // Цитология. — 1979. — T. 21, No 5. — С . 508–513.
- Балушкина Е. В., Петрова Н.А. Функционировние популяций хирономид в гипергалинных озерах Крыма // Тр. ин-та Зоол. ин-т. —Л.: Наука, Т. 205. – 1989. — С. 129–138.
- Белянина С.И. Кариотипический анализ хирономид (Chironomidae, Diptera) фауны СССР: дис…. докт. биол. наук. – М. – 1983. 418 с.
- Белянина С.И. Современное состояние кариофондов хирономид в водоемах СССР // Эволюция, видообразование и систематика хирономид. Новосибирск: изд-во Новосиб. ун-та. 1986. С. 45–49.
- Белянина С. И. Сравнительный хромосомный анализ популяций фитофильной хирономиды Glyptotendipes glaucus (Mg.) затронутых чернобыльским выбросом // Генетика. — 2014а. — Т. 50, №9. — С. 1025-1032.
- Белянина С.И. Современное состояние кариофондов хирономид (Chironomidae, Diptera) в водоемах Брянской области // Радиобиология: антропогенные излучения: Матер. междунар. науч. конференции Гомель, 2014б. — С. 12-14.
- Белянина С.И. Состояние хромосом и кариофондов Chironomusplumosus (L.) (Chironomidae, Diptera) на затронутых Чернобыльским выбросом территориях // Экология и защита окружающей среды: Сборник Междунар. научно-практ. конференции. — 2014 в. — С. 70-72.
- Белянина С.И., Кузьмина К.А., Сигарева Л.Е. Гигантские хромосомы хирономид как модель для изучения генетических эффектов пестицидов на водные организмы // Гигиенические аспекты охраны окружающей среды: Научно-тематический сб. Саратов: Изд-во Сарат. ун-та, — 1979. — С. 80-83.
- Белянина С.И., Колосова Т.А. Кариотип Chironomus behningi из бассейна Аральского моря // Цитология.-1979.- Т. 29. №9.- С. 1103–1106.
- Белянина С.И., Мосияш С.А. Кариофонд природной популяции Chironomusplumosus из Иваньковского водохранилища в районе Конаковской ГРЭС // Новые данные по кариосистематике двукрылых насекомых. – Л.: ЗИН АН СССР. - 1980. - С. 12–15.
- Белянина С.И., Гаврикова В.В., Логинова Н.В. (Полуконова). Влияние ряда ксенобиотиков на генетическую активность и структуру гигантских хромосом хирономид // Экология и здоровье: Тез.докл. науч-практ. конф. Саратов: СГМУ. 1993. – С. 52–53.
- Белянина С.И., Логинова Н.В. (Полуконова), Сигарева Л.Е. Морфологическая характеристика вида Chironomusentis (Chironomidae, Diptera) и его кариофонд из разных географических зон // Зоол. журн. – 1992. – Т. 71, №8. - С. 32–38.
- Влияние ботулинического токсина на генетическую активность гигантских хромосом. С.И. Белянина, Н.П. Чеснокова, А.О. Колбенев, Т.А. Невважа// Бактериально-вирусные инфекции. Ч. 1. Саратов: СГМУ, 1993.– С. 90–94.
- Белянина С.И., Воронин М.Ю., Белоногова Ю.В. Влияние искусственного повышения температуры в водоеме-охладителе Балаковской АЭС на структурно-функциональное состояние хромосом хирономид (Chironomidae, Diptera) // В кн.: Биоразнообразие и устойчивость живых систем. Матер. XIII Междунар. научно-практ. экологической конференции. (г. Белгород, 6-11 октября 2014 г.). –2014. –С. 119-120.
- Бецкий О.В., Кислов В.В., Лебедева Н.Н. Миллиметровые волны и живые системы. – М.: Сайнс пресс. – 2004. – 271 с.
- Благой Ю.П. Взаимодействие ДНК с биологически активными веществами (ионами металлов, красителями, лекарствами) // Соросовский Образовательный Журнал. – 1998. – Т. 10. – С. 18-24.
- Влияние низкоинтенсивного КВЧ - излучения на генетическую активность политенных хромосом Chironomusplumosus. Г.Е. Брилль, О.Р. Апина, С.И. Белянина, Н.П. Панина // Физическая медицина. – 1993. – Т. 3, №1–2. – С. 71.
- Бухтеева Н.М. Реакция ядерного аппарата клеток слюнных желез личинок 4-го возраста и предкуколок Camptochironomustentans на воздействие глинозема // Морфология и морфогенез тканей и органов мезенхимного происхождения. Иркутск: изд-во Иркутск. ун-та. 1975. С. 7–9.
- Валеева Ф.С. Действие высоких доз актиновмицина на структуру, пуффинг и транскрипцию политенных хромосом слюнных желез Chironomusthummi // Цитология. – 1979. –Т. 21, № 12. – С. 1411-1418.
- Василенко О. И. Радиационная экология. - Татарстан: Изд-во «Медицина». 2004. – 215 с.
- Величко А. К. Влияние теплового шока на репликацию ДНК, стабильность генома и структуру хроматина: Автореф. дис. канд. биол. наук. – М., 2012. – 28 с.
- Гиляров М. С. Биологический энциклопедический словарь. – М.: Сов. Энциклопедия, 1986.
- Голосова А.В., Пак И.В., Кузнецова Т.Ю. Генотоксические эффекты пестицидов: дельтаметрина (дециса) и метсульфуронметила (магнума) // Вестник экологии, лесоведения и ландшафтоведения. Тюмень: ИПОС СО РАН. – 2009. Вып. 10. – С. 101-108.
- Молекулярно-цитогенетическая диагностика сложных хромосомных перестроек при остром миелоидном лейкозе. М.М. Дегтярева, Т.В. Карамышева, О.В. Саблина, Н.Б. Рубцов // Клиническая онкология и гематология. – 2000. – № 2. – С. 22-25.
- Демин С.Ю. Изменчивость степени конденсированности политенных хромосом в клетках разных органов личинок Chironomusplumosus из природы. Автореф. дис. … канд. биол. наук. – Л., 1989. – 25 с.
- Демин С.Ю., Ильинская Н.Б. Изменение компактности политенных хромосом из разных органов личинок мотыля Chironomusplumosus // Цитология. – 1988. – Т. 30, №4. – С. 407– 415.
- Дроздовская Л.Н. Влияние бора на развитие пуфов в политенных хромосомах слюнных желез Drosophilamelanogaster на стадии ранней предкуколки // Генетика. — 1974. — Т. 10, №5. — С. 57–64.
- Влияние 5-фторурацила на спектр пуфов политенных хромосом слюнных желез Drosophilamelanogaster. Л.Н. Дроздовская, И.А. Рапопорт, М.М. Бекназарьянц, Н.М. Вишнякова //Эффективность химических мутагенов в селекции. М.: Наука. — 1976. — С. 73–85.
- Дроздовская Л.Н., Бекназерьянц М.М., Рапопорт И.А. Химический мутагенез и создание сортов интенсивного типа. – М.: Наука, 1977. – С. 212–219.
- О генетических процессах в популяциях, подвергающихся хроническому воздействию ионизирующей радиации. Н.П. Дубинин, В.А. Шевченко, А.А. Алексеенок, и др.// Успехи совр. генетики. – 1972. – №4. – С. 170–205.
- Дубровский Э.Б., Козлова Т.Ю., Ржецкая М.А. Регуляция онтогенетической экспрессии генов теплового шока из пуфа 67В в слюнных железах Drosophilamelanogaster // Онтогенез.-1990.–Т.21– №5.–С. 508–516.
- Дурнова Н.А., Сигарева Л.Е., Оглезнева А.А. Пространственная организация политенных хромосом в интерфазных ядрах периферических хирономид (Diptera, Chironomidae) // Труды Русского энтомологического общества. СПб., 2014. – Т. 85(2). – 27-39.
- Жимулев И.Ф. Политенные хромосомы: морфология и структура. Новосибирск. «Наука». – 1992. – 479 с.
- Жимулев И.Ф. Гетерохроматин и эффект положения гена. Н.: ВО Наука. Сиб. изд. фирма. – 1993. – 491 с.
- Жимулев И. Ф. Современные представления об организации и функционировании политенных хромосом // Соросовский Образовательный Журнал. 1997. – Т. 11. – С. 2-7.
- Жимулев И.Ф., Беляева Е.С. К вопросу о структурно-функциональной организации политенных хромосом // Генетика. – 1975. – Т. 11. – №2. – С. 175–181.
- Жимулев И.Ф., Беляева Е.С. Изменение структуры политенных хромосом дрозофилы при длительном культивировании слюнных желез личинок в брюшной полости взрослых мух // Цитология. – 1976. – Т. 18, № 1. – С. 5–11.
- Жимулев И.Ф., Куличков В.А. Районы разрывов политенных хромосом Drosophilamelanogaster: локализация и особенности репликации // Генетика. – 1977. – Т. 13. № 1. – С. 85–94.
- Зайниев Г.А. Кольцо Бальбиани – симметрично организованный пуффф, «выплеснувшийся» из хромосомы // Структура и функции клеточного ядра. 9 Всем. Симп. Черноголовка, – 1987. – С. 56.
- Ильинская Н.Б. Изменение политенных хромосом в сезонном цикле развития Chironomusplumosus L. (Diptera, Chironomidae) и их значение для кариосистематики // Экологические и морфологические основы систематики двукрылых насекомых. – Л.: изд-во ЛГУ, 1979. – С. 29–39.
- Ильинская Н.Б. Морфологическая изменчивость политенных хромосом личинок хирономид в естественных условиях обитания. Автореф. дисс. … докт. биол. наук. Л., 1989. 38 с.
- Ильинская Н.Б., Максимова Ф.Л. Изменчивость политенных хромосом слюнных желез личинок Chironomusplumosus в разные месяцы года // Цитология. – 1976. – Т. 18, – №6. – С. 747–751.
- Ильинская Н.Б., Максимова Ф.Л. Изменение рисунка и числа дисков в политенных хромосомах Chironomusplumosus в разные сезоны года // Цитология. – 1978. – Т. 20, – №3. – С. 291–296.
- Кармоков М.Х., Хатухов А.М., Полуконова Н.В. Кариотипическая структура двух видов рода Chironomus (Chironomidae, Diptera) Центрального Кавказа // Труды Ставропольского отделения Русского энтомологического общества. Вып. 4: материалы Международной научно-практической конференции. – Ставрополь: АГРУС, – 2008. – С. 255-257.
- Кикнадзе И.И. Анализ работы пуффов в ходе личиночного развития двукрылых на фоне экспериментальных воздействий // Клеточная дифференцировка и индукционные механизмы. М. Наука, 1965. – С. 78-82.
- Кикнадзе И.И. Политенные хромосомы как модель интерфазной хромосомы // Цитология. – 1971. — T. 13. – N 6. — С. 716–732.
- Кикнадзе И.И. Функциональная организация хромосом. – М., Л.: Наука, 1972. – 211 с.
- Кикнадзе И.И. Молекулярно-цитологическая организация колец Бальбиани и генов, локализованных в них // Организация и экспрессия генов тканеспецифической функции у Diptera. – Новосибирск: Наука. – 1985–. С. 23–97.
- Кикнадзе И.И., Беляева Е.С. Ядрышко, закономерности его форми-рования и генетическая роль // Генетика. – 1967. – Т. 7, N 8. – C. 149–161.
- Изменение массы прицентромерного гетерохроматина – один из важных путей эволюции кариотипа у хирономид. И.И. Кикнадзе, М.Т. Сиирин, М.А. Филиппова, и др. // Цитология.–1991.– №2. – С. 90–98.
- И.И. Кикнадзе, А.Г. Истомина, Л.И. Гундерина и др. Кариофонды хирономид криолитозоны Якутии: Триба Chironomini. Новосибирск: Наука, 1996. – 166 с.
- Кариофонд голарктической хирономиды Glyptotendipesbarbipes (Staeger). И.И. Кикнадзе, Е.Н. Андреева, А.Г. Истомина, М.Дж. Батлер // Цитология. — 1998. – Т. 40. – No10. – С. 900–912.
- Кикнадзе И.И., Истомина А.Г. Кариотипы и хромосомный полиморфизм сибирских видов хирономид (Diptera, Chironomidae) // Сиб. экол. журн. — 2000. — №4. — С. 445–460.
- Куберская Е.Ф., Бухтеева Н.М. Некоторые реакции личинок хирономид на воздействие фенола различных концентраций // Пато- и саногенетические реакции на различных уровнях организма. – Иркутск: Изд-во Иркутск. ун-та, 1975. – С. 41–43.
- Кузин А.М. Роль природного радиационного фона и вторичного биогенного излучения в явлении жизни. М.: Наука. — 2002 г. — 79 с.
- Куринный А. И. К проблеме предупреждения генетических последствий применения пестицидов: реальность и необходимость // Цитология и генетика. – 1983. – № 7. – С. 16–21.
- Лычев В.А. Изучение активности хромосом при глубоком инбридинге у дрозофилы // Цитология. – 1965. Т. 7. – №3. – С. 325–333.
- Лычёв В. А. Изучение величины и распределения пуфов Drosophila melanogaster в норме и при влиянии инбридинга и облучения: Дис. канд. биол. наук. Обнинск. 1968. 202 с.
- Заседателев А.С, Гурский Г.В., Готтих Б. П. Аналоги актиномицина D с заместителями в положении 7 феноксазонового хромофора. СМ. Никитин, С.Л. Гроховский, А.Л. Жузе, М.В. Михайлов // Биоорганическая химия. — 1981. — Т.7, № 4. – С. 542-551.
- Петрова Н.А. Хромосомные перестройки трех видов хирономид из зоны Чернобыля (Diptera, Chironomidae) // Генетика. – 1991. – Т.27, №5. – С. 836–848.
- Петрова Н.А., Максимова Ф.Л. Роль хромосомных перестроек в видообразовании хирономид // Генетика. — 1978. — Т. 14, №7. — С. 1201–1207.
- Полуконова Н.В. Показатели хромосомного полиморфизма хирономид на антропогенно-трансформированных территориях // Новые технологии в защите биоразнообразия в водных экосистемах: Междунар. конфер.: Москва, 27-29 мая 2002 г.: Тезисы докл. М.: МАКС Пресс. — 2002. — С. 85.
- Полуконова Н.В., Сокова А. В, Козлов М.С. Клеточные механизмы адаптации хирономид в условиях длительного воздействия низких температур // Окружающая среда и гигиена. Мат. науч.-практ. конфер. Саратов: Изд-во СГМУ. — 2002. — С. 78–80.
- Полуконова Н.В., Филинкова Т.Н., Сокова А.В. Эколого-кариологический подход к исследованию популяций хирономид севера // Проблемы экологии и биоразнообразия. Тез. докл. межд. конф. мол. уч. Борок, ИБВВ. – 2002. – С. 69.
- Полуконова Н.В., Белянина С.И. О возможности гибридогенеза в видообразовании комара-звонца Chironomususenicus Loginova et Beljanina (Chironomidae, Diptera) // Генетика. – 2002. – Т. 38, №12. – С. 1635–1640. Polukonova N.V, Beljanina S.I. On the Possibility of Hybridogenesis in the Origin of Midge Chironomus usenicus Loginova et Beljanina (Chironomidae, Diptera) // General Genetics. – 2002. – 12. – S. 1385–1390.
- Полуконова Н.В., Белянина С.И. Солоновато-водные хирономиды (Diptera) Каспийского моря и его бассейна: особенности морфологии, филогенетические связи и эволюция // Эколого-биологические проблемы бассейна Каспийского моря: Мат. Всерос. науч. конф. Астрахань: Изд-во Астраханского гос. ун-та. – 2003. – С. 97.
- Полуконова Н.В., Белянина С.И., Гурина О.В., Воронин М. Особенности структуры кариотипа Lipiniellaarenicola Shilova из водоемов Саратовской области (Diptera, Chironomidae) // Новые данные по хирономидологии. Борок: ИБВВ РАН. — 2003а. — С. 1-9.
- Полуконова Н.В., Воронин М.Ю., Ермохин М.В. Хромосомный полиморфизм личинок хирономид и возможности хромосомного мониторинга экосистемы водоема-охладителя Балаковской АЭС // Тезисы докладов международной и Молодежной конф., Россия, Тольятти, 15-19 сентября 2003 г. Тольятти: ИЭВБ РАН. – 2003б. – С. 226.
- Полуконова Н.В., Белянина С.И., Михайлова П.В., Голыгина В.В. Сравнительный анализ кариотипов и кариофондов комаров-звонцов Chironomusnuditarsis и Ch. curabilis (Chironomidae, Diptera) // Зоол. журн. — 2005. — Т. 84, №2. — С. 195–206.
- Полуконова Н.В., Филинкова Т.Н. Морфология политенных хромосом и особенности инверсионного полиморфизма хирономин (Chironomidae, Diptera) севера России // Энтомологические и паразитологические исследования в Поволжье: Сб.науч.тр. Вып. 5. Саратов: Изд-во Сарат.ун-та, – 2006. — С. 18-21.
- Полуконова Н.В., Белянина С.И. Цитогенетические эффекты ксенобиотиков у Chironomus riparius (Chironomidae, Diptera) // Саратовский научно-медицинский журнал.– 2006. — № 3. — С. 33-39.
- Полуконова Н.В., Фёдорова И.А. Эколого-кариологическая оценка последствий действия экологических факторов на хирономид (Chironomidae, Diptera) // Поволжский экологический журн. — 2006. — № 2/3. — С.164-175.
- Полуконова Н.В., Воронин М.Ю. Особенности хромосомного полиморфизма и транскрипционной активности политенных хромосом массовых видов хирономид водоема-охладителя Балаковской АЭС // Энтомологические и паразитологические исследования в Поволжье: Сб.науч.тр. Саратов: Изд-во Сарат.ун-та. – 2006. Вып. 5. — С. 67-69.
- Полуконова Н.В., Белянина С.И., Фёдорова И.А. Методологические подходы к анализу генотоксичности // XXI Любищевские чтения. Современные проблемы эволюции (сборник докладов). Ульяновск: Ульяновский гос. педуниверситет. — 2007а. — С. 180-186.
- Анализ сообществ, популяций и генома хирономид (Diptera) в биомониторинге гидроэкосистем. Н.В. Полуконова, А.Г. Дёмин, И.В. Катаева, И.А. Фёдорова // «Водные экосистемы, организмы, инновации – 9»Тез. 9-й конференции (сессии стендовых сообщений) М.: МГУ, 2007 б. – С. – 102.
- Биологический мониторинг водных экосистем на основе анализа сообществ, популяций, кариотипа и мтДНК хирономид (Chironomidae, Diptera). Н.В. Полуконова, М.В. Ермохин, М.Ю. Воронин, и др. // Известия Саратовского гос.университета. Новая серия. Т. 7. Серии Химия. Биология. Экология, 2007в. — С. 72-81.
- Полуконова Н. В., Демина И. В., Демин А. Г., Кармоков М. Х., Федорова И. А. Возможности и перспективы использования комаров-звонцов (Chironomidae, Diptera) при комплексном подходе в биологическом мониторинге водных экосистем и токсикологических исследования // Материалы IV всероссийского симпозиума по амфибиотическим насекомым и X трихоптерологического симпозиума «Проблемы водной энтомологии России и сопредельных государств». СОГУ. — 2010. — С. 69-73.
- Полуконова Н.В., Кармоков М.Х. Микроэволюционные изменения в популяциях Chironomus nuditarsis Str. (Keyl, 1962) (Chironomidae, Diptera) Центрального Кавказа // Генетика. — 2013. — Т. 49. №2. — С. 1-7.
- Полуконова Н.В., Курчатова М.Н., Дурнова Н.А. Применение нового тест-объекта личинок Chironomusriparius для оценки токсичности на примере экстракта Gratiolajficinalis // Токсикологический вестник. — 2014. — С. 334-345.
- Полякова А.Г. КВЧ- и лазеропунктура в комплексной медицинской реабилитации пациентов с патологией суставов и позвоночника: (клинико-эксперим. исслед.): автореф. дис. … д-ра мед. наук. – М, 2004. – 47 с.
- Полякова А.Г. Механизмы влияния низкоинтенсивного электромагнитного излучения крайне высокочастотного и терагерцевого диапазонов на процессы репаративной регенерации соединительной ткани // Успехи современного естествознания. – 2013. – № 4 . – С. 32-36.
- Прокофьева-Бельговская А.А. Организация и функционирование хромосом // Вестник. АН СССР. — 1962. — № 12. — С. 60-62.
- Прокофьева-Бельговская А.А. Гетерохроматинные районы хромосом: строение и функции // Ж. общ. биол. — 1977. — Т. 38. – №5. — С. 735–741.
- Розен В. Б. Основы эндокринологии. — 3-е изд., перераб. и доп. — М.: Изд. МГУ: Медицина, 1994. — С. 40—93. — 384 с.
- Рубцов Н. Б., Карамышева Т. В. Системы идентификации гомологичных районов хромосом: сравнительная цитогенетика млекопитающих и хромосомные патологии человека. — Новосибирск. 1996. — 183 с.
- Рубцов Н.Б., Карамышева Т.В. Цитогенетическая диагностика онкологических заболеваний. // Клиническая онкология и гематология. — 2000. — Т.2. — С.7-21.
- Обратная in situ гибридизация ДНК-зондов аномальных хромосом в диагностике хромосомных патологий. Н.Б. Рубцов, Т.В. Карамышева, В.Г Матвеева., О.В. Саблина и др. // Генетика. — 2001. — Т. 37. – №10. — С. 1-8.
- Сахарова М.Н., Рапопорт И.А. Пуфы, индуцированные роданидом и пуфовая модель определения поражаемых лекарствами ферментов // Докл. АН СССР. — 1971. — Т. 196.– №5. — С. 1217-1270.
- Сигарева Л.Е. Изучение структурного гетерохроматина в дифференциально окрашенных политенных хромосомах хирономид. Автореф. дисс. … канд. биол. наук. – Саратов, 1985. 197 с.
- Иммунный ответ при вирусных инфекциях. Т.В. Сологуб, М.Ю. Ледванов, В.П. Малый и др. // Успехи современного естествознания. – 2009. – № 12. – С. 29-33.
- Соколова Н.Ю. (ред.) Мотыль Chironomusplumosus L. (Diptera, Chironomidae). Систематика, морфология, экология, продукция. – М, 1983. — 307 с.
- Фёдорова И. А. Характер изменения функционально активных участков и компактности политенных хромосом Chironomus (Diptera) под влиянием холинотропных препаратов: Авторефэ дис. канд. биол. наук. – Астрахань, 2008. – 19 с.
- Фёдорова И.А., Полуконова Н.В. Оптимизация методов хромосомного мониторинга водных экосистем разного типа // Экологические проблемы урбанизированных территорий: материалы научн.-практ. конференции. Елец: ЕГУ им. И.А. Бунина, 2007а. – С. 151-155.
- Фёдорова И.А., Полуконова Н.В. Особенности цитогенетического анализа ткани слюнных желез личинок хирономид под воздействием холинотропных препаратов // Тез. Всероссийской научной конференции. Астрахань, 2007б. – С. 191.
- Фёдорова И.А., Полуконова Н.В. Цитогенетические эффекты политенных хромосом хирономид под влиянием холинотропных препаратов // Биология внутренних вод: –Тез. докл. XIII международной школы-конференции молодых учёных. Борок, – 2007 в. – С. 54.
- Фёдорова И.А., Полуконова Н.В. Реакция различных участков генома хирономид (Diptera) под действием атропина // Энтомологические и паразитологические исследования в Поволжье. Саратов. Изд-во Сарат. ун-та. — 2007 г. — Вып. 6. — С. 65-73.
- И.А. Фёдорова, Е.Н. Балалаева, Ю.В. Журавлёв, Е.Л. Бодякина Принципы оценки генетической активности политенных хромосом хирономид (Chironomidae, Diptera) // Ломоносов: Тез. докл. международной научной конференции студентов, аспирантов и молодых ученых. М.; МГУ, 2007. – С. 25.
- Фёдорова И.А., Полуконова Н.В. Изменение транскрипционной активности политенных хромосом Chironomusplumosus (Diptera) под действием полулетальной концентрации атропина // Труды Ставропольского отделения Русского энтомологического общества. Вып. 4: материалы междунар. научно-практ. конфер: Ставропольский гос. аграрный университет. – Ставрополь: АРГУС. – 2008. – С. 269-273.
- Федорова И.А., Полуконова Н.В., Петров Н.В. Цитогенетические эффекты холинотропных препаратов при комбинированном действии на личинок Chironomus рlumosus (Diptera) in vivo // Цитология. — 2009а. — Т. 51, № 10. — С. 849-857.
- Функциональная активность политенных хромосом Chironomus (Diptera) под влиянием холинотропных препаратов атропина и пилокарпина. И.А. Фёдорова, Н.В. Полуконова, К.Н. Дворецкий, С.И. Богословская // Экологическая генетика. — 2009б. — Т.7. – №3. — С. 44-52
- Филинкова Т.Н. Хирономиды родов Chironomus Meigen и Camptochironomus Kieffer (Diptera, Chironomidae) Полярного Урала, Ямала и Приобской лесотундры (морфология и кариотипические особенности. Автореф. дис. … канд. биол. наук. – СПб, 1995. – 16 с.
- Христолюбова Н.Б. Образование дополнительных ядрышек в клетках слюнных желез дрозофил под действием трипсина // Морфологические и химические изменения в процессе развития клетки. – Рига: Зинатне, 1968. – С.95–99.
- Чуян Е.Н., Раваева М.Ю., Трибрат Н.С. Низкоинтенсивное электромагнитное излучение миллиметрового диапазона: влияние на процессы микроциркуляции // Физика живого. – 2008. – Т.16. – № 1. – С. 82–90.
- Шобанов Н.А. Кариофонд Chironomusplumosus (L.) (Diptera, Chironomidae). IV: Внутри- и межпопуляционный полиморфизм // Цитология. — 1994. — Т. 36, №11. — С. 1129–1145.
- Шобанов Н.А. Эволюция рода Chironomus (Diptera, Chironomidae). 1. Предковая форма и основные направления филогенеза // Зоол. журн. – 2002а. – Т. 81. №4. – С. 463-468.
- Шобанов Н.А. Эволюция рода Chironomus (Diptera, Chironomidae). 2. Филогенетическая модель // Зоол. журн. – 2002б. – Т. 81. – №6. — С. 711-718.
- Шобанов Н.А., Зотов С.Д. Цитогенетические аспекты филогении рода Chironomus Meigen (Diptera, Chironomidae) // Энтомол. обозр. – 2001. – Т. 53, No1. – C. 180–193.
- Ямполь Г.П. Временное напряжение контакта политенных хромосом с ядерной оболочкой как отражение перестройки транскрипционной активности генома в результате стрессорных физиологических и экспериментальных воздействий // Тез. докл. Всесоюз. об-ва генетиков и селекционеров, — 1987. — С. 315.
- Ярмоленко С. П., Вайнсон А. А. Радиобиология человека и животных. – М.: Высш.шк. 2004. – 548 с.
- Buck-Koehntop, B. A.; Defossez, P.-A. On how Mammalian Transcription Factors Recognize Methylated DNA. – Epigenetics, – 2013. – 8. – 131-137.
- Beermann W. Riesenchromosomen // Protoplasmatologica. – 1962. – Vol. 6. – S. 1–161.
- Berendes H.D. Synthetic activity of polytene chromosomes // Intern. Rev. Cytol. — 1973. — Vol. 35. — P. 61–116.
- Bethany A., Pierre-Antoine. On how mammalian transcription factors recognize methylated DNA // Epigenetics. — 2013. — 8(2). — Р. 131—137.
- Blaylok B. G. Chromosomal polymorphis in irradiated natural populatias of Chironomus // Genetics. — 1966. — Vol. 53, №1. — P. 131–136.
- Boyd J. B., Boyd S.F. Influence of BDU on the stability and function of polytene chromosomes // Chromosoma. — 1977. — Vol. 61, №1. — P. 75–94.
- Foti James J., Devadoss Babho, Winkler Jonathan A., Collins James J., Walker Graham C. Oxidation of the guanine nucleotide pool underlies cell death by bactericidal antibiotics // Science. 2012. V. 336. Pp. 315–319.
- Gratzner H. G. Monoclonal antibody to 5-bromo-and 5-iododeoxyuridine: a new reagent for detection of DNA replication // Science. – 1982. – Т. 218. – №. 4571. – С. 474-475.
- Chakrabarti R., Schutt C.E. The enhancement of PCR amplification by low molecular-weight sulfones» // Gene. — 2001. — Т. 274 (1-2). — 293–298.
- Clever U. Genaktivitalen in den Riesenchromosomen von Chironomus tentans und Beziehungen zur Entwicklung. III Das Aktivitatis – muster in Phasen der Entwicklung – Sruhe // J. Insect. Physiol. — 1962. — №2. — P. 357–376.
- Hägele K. DNS – Replication muster der Speicheldrüsenchromosomen von Chironomiden // Chromosoma. — 1970. — Vol. 31, №1. — P. 91–138.
- Heyn H, Sayols S, Moutinho C.. Esteller M. et al. Linkage of DNA Methylation Quantitative Trait Loci to Human Cancer Risk // Cell Reports. — 2014. — 7. Р. 331–338.
- Hodgson, D. A. Primary metabolism and its control in Streptomycetes. Adv. Microb. Physiol. — 2000. — 42. 4— 7-238.
- Ineichen H. Photoperiodsche Kontrolle der Entwicklung von Chironomus tentans und Entwicklungsspecifishe Puff Veranderungen (in vivo und in vitro) // Rev. Suisse. Zool. — 1978. — Bd.85. — S. 87–89.
- Keyl H.G. Chromosomenevolution bei Chironomus II. Chromosomenbauten und phylogenetische Bezichungen der Arten // Chromosoma. — 1962. — Vol. 13, №4. — S. 464-514.
- Michailova P.V. Untersuchungen uber den Chromosomen polymorphisms bei Chironomus salinarius Kieff., Chironomus valkanovi Michailova und Chironomus anchiatus Michailova (Diptera, Chironomidae) von der bulgarischen Schwartzmeerkuste // Zool. Anz. Leipzig. — 1973. — Bd. 191. — S. 348–364.
- Michailova P.V. Structural changes in the polytene chromosomes of Chironomus valkanovi Michailova, Cricotopus silvestris Staeg. and Cricotopus varius Fabr. (Dipreta, Chironomidae) under the influence of various factors // Comp. Rend. Acad. Bulg. Sci. — 1979. — Vol. 32. — S. 377–380.
- Michailova P. The politene chromosomes and their significance to the systematics and phylogeny of the family Chironomidae, Diptera // Acta Zool. Fenn. Suppl. — 1989. — 107 p.
- Michailova P. V. Karyosystematics and morphology of Chironomus salinarius type larvae (Diptera, Chironomidae) from Finland // Cytobios. — 1992. — 71. — P.163-169.
- Chromosome rearrangements, repetitive DNA clusters and tpansposable elements in Chironomus riparius Mg. (Diptera, Chironomidae) from anthropogenically polluted regions. P. Michailova, J. Ilkova, Sella et al. // Book of adstracts IV international conference on karyosystematics of the invertebrates. St. Petersburg: Nauka, 2006. – P. 38.
- Mitelman F., Mertens F, Johansson B. A breakpoint map of recurrent chromosomal rearrngements in human neoplasia // Nature Genetics. Special issue. — 1997. — P.415-474.
- Potter A.M., Watmore A. Cytogenetics in myeloid leukaemia // Human Cytogenetics: a practical approach. — 1992. — V.1 1. — P. 27-67.
- Rabbitts TH. Chromosomal translocations in human cancer // Nature. — 1994. — V. 372. — P. 143-149.
- Sass M., Heinz L. Hierarchy of fibrillar organization levels in the polytene interphase chromosomes of Chironomus // J. Cell Sci. — 1980. — Vol. 45. — P. 269–293.
- Serfling E., Panitz R., Wobus U. Die experimentelle Beeinflussung des puffmusters von Riesenchromosomen. Puffinduktion durch Oxytetracyclin bei Chironomus thummithummi // Chromosoma. — 1969. — 28. № 2. — 107–119.
- Widmer RM, Lucchini R, Lezzi M, Meyer B, Sogo JM, Edstr6m J-E, Koller T (1984) Chromatin structure of a hyperactive secretory protein gene (in Balbiani ring 2) of Chironomus. EMBO J 3:1635-1641.
- Measurement of cell kinetics in human tumours in vivo using bromodeoxyuridine incorporation and flow cytometry. G. D. Wilson et al. // British journal of cancer. – 1988. – Т. 58. – №. 4. – С. 423.
- Wulker W., Butler M.G. Karyosystematics and morphology of northern Chironomus (Diptera, Chironomidae): Freshwater species with larvae of the salinarius-type // Entomol. Scand. — 1983. — Vol. 14, №1. — P. 121–136.
Таблицы
Таблица 1.Морфофункциональные особенности функционально активных участков ПХ
|
Показатели (авторы описания) |
Особенности морфологии ПХ |
|
|
При увеличении функциональной активности |
При уменьшении функциональной активности |
|
|
Состояние хромомера [35, 73, 74, 77, 101, 102] |
Декомпактизация материала хромомера и образование пуфа |
Увеличивается четкость хромомера, вызванная компактизацией материала хромомера |
|
ЯО [45, 49, 73, 74, 77 101, 102] |
Увеличение размеров |
Уменьшение размеров, появление четкого дискового рисунка |
|
Пуфы [73, 74, 77, 101, 102] |
Активность крупных пуфов изменяется стадиеспецифично. Малые пуфы в основном активны на протяжении всего развития |
Уменьшение размеров |
|
КБ [73. 74, 77, 101, 102, 137] |
Увеличение размеров. При обработке пилокарпином – до 370 молекул РНК-полимеразы на ген |
Уменьшение размеров. |
|
Длина хромосомы [73, 74, 77, 101, 102] |
Увеличение |
Уменьшение |
|
Способность гомологов ПХ к конъюгации [74] |
Не изменяется |
Ослабление приводит к формированию «помпонообразной» хромосомы |
|
Состояние междисков [37, 74, 101, 102] |
Хорошо выражены |
Материал междисков компактизируется, и рядом расположенные диски объединяются в блоки неактивного хроматина – образуются «новые» диски |
Таблица 2.Метрические показатели и индексы ФА ПХ на примере Chironomusplumosus
|
Обоз-наче-ние |
Название индекса |
Расчет индекса |
|
К/Е |
Индекс компактности хромосомы III |
Отношение абсолютной длины плеча Е к ширине центромеры |
|
Кn |
Коэффициент ФА ЯО хромосомы IV |
Отношение максимального диаметра ядрышка к ширине интактного района 6 хромосомы IV |
|
А/d1G* |
Коэффициент ФА КБ1 хромосомы IV |
Отношение максимального диаметра КБ1 к ширине интактного района 6 хромосомы IV |
|
А/d2G * |
Коэффициент ФА КБ2 хромосомы IV |
Отношение максимального диаметра КБ2 к ширине интактного района 6 хромосомы IV |
|
А/dВ* |
Коэффициент ФА КБ плеча В хромосомы I |
Отношение максимального диаметра КБ плеча В к ширине интактного района 17 того же плеча хромосомы I |
|
Pв* |
Коэффициент ФА пуфа плеча В хромосомы I |
Отношение максимального диаметра пуфа 21 района плеча В к ширине интактного диска 21 |
*Примечание: звездочкой отмечены индексы, обозначения для которых предложены нами [101, 102]
